Grāmata:
Ūdens blīvums- Tas ir faktors, kas nosaka ūdens organismu pārvietošanās apstākļus un spiedienu dažādos dziļumos. Destilētam ūdenim blīvums ir 1 g / cm 3 4 ° C temperatūrā. Dabisko ūdeņu, kas satur izšķīdušos sāļus, blīvums var būt lielāks, līdz 1,35 g / cm 3. Spiediens palielinās līdz ar dziļumu par aptuveni 1 · 10 5 Pa (1 atm) vidēji uz katriem 10 m.
Krasā spiediena gradienta dēļ ūdenstilpēs ūdens organismi parasti ir daudz eirobātiskāki nekā sauszemes organismi. Dažas sugas, kas izplatītas dažādos dziļumos, pieļauj spiedienu no vairākiem līdz simtiem atmosfēru. Piemēram, Elpidia ģints jūras gurķi, Priapulus caudatus tārpi apdzīvo no piekrastes zonas līdz ultraabyssal. Pat saldūdens iemītnieki, piemēram, skropstiņi, čības, suvoji, peldvaboles utt., eksperimentā var izturēt līdz 6 · 10 7 Pa (600 atm).
Tomēr daudzi jūru un okeānu iedzīvotāji ir relatīvi stenobāti un atrodas noteiktā dziļumā. Stenobātisms visbiežāk ir raksturīgs sekla un dziļūdens sugām. Tikai piekrastes joslu apdzīvo annelidsmilšu tārps Arenicola un jūras šķīvīši (Patella). Daudzas zivis, piemēram, no makšķernieku grupas, galvkāji, vēžveidīgie, pogonofori, jūras zvaigznes u.c. sastopamas tikai lielā dziļumā vismaz 4 · 10 7 - 5 · 10 7 Pa (400-500 atm) spiedienā.
Ūdens blīvums nodrošina iespēju uz to paļauties, kas ir īpaši svarīgi skeleta formām. Vides blīvums kalpo kā nosacījums planēšanai ūdenī, un daudzi ūdens organismi ir pielāgoti šim konkrētajam dzīvesveidam. Suspendētie, ūdenī peldošie organismi tiek apvienoti īpašā ekoloģiskā ūdens organismu grupā - planktons ("Planktos" - planējošs).
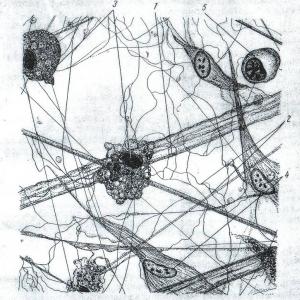
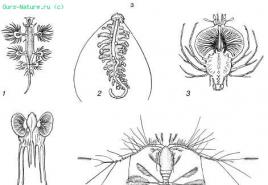
Rīsi. 39. Ķermeņa relatīvās virsmas palielināšanās planktona organismos (saskaņā ar S. A. Zernovu, 1949):
A — stieņa formas:
1 - diatoma Synedra;
2 - zilaļģes Aphanizomenon;
3 - Peridineal aļģes Amphisolenia;
4 - Euglena acus;
5 - galvkāji Doratopsis vermicularis;
6 - copepod Setella;
7 - Porcelānas kāpurs (Decapoda)
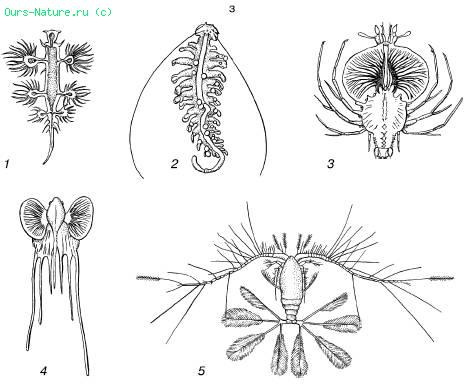
B — sadalītas formas:
1 - gliemji Glaucus atlanticus;
2 - tārps Tomopetris euchaeta;
3 - vēžu Palinurus kāpurs;
4 - jūrasvelna Lophius zivju kāpurs;
5 - copepod Calocalanus pavo
Planktonā ir vienšūnu un koloniālās aļģes, vienšūņi, medūzas, sifonofori, ķemmes želejas, pterigopodi un gliemenes, dažādi mazie vēžveidīgie, bentosa dzīvnieku kāpuri, zivju ikri un mazuļi un daudzi citi (39. att.). Planktona organismiem ir daudz līdzīgu pielāgojumu, kas palielina to peldspēju un neļauj tiem nogrimt dibenā. Šādi pielāgojumi ietver: 1) vispārēju ķermeņa relatīvās virsmas palielināšanos izmēru samazināšanās, saplacināšanas, pagarinājuma, daudzu izaugumu vai saru veidošanās dēļ, kas palielina berzi pret ūdeni; 2) blīvuma samazināšanās sakarā ar skeleta samazināšanos, organismā uzkrājoties taukiem, gāzes burbuļiem u.c.. Kramaļmatos rezerves vielas nogulsnējas nevis smagās cietes, bet tauku pilienu veidā. Nightlight Noctiluca izceļas ar tādu gāzu vakuolu un tauku pilienu pārpilnību šūnā, ka tajā esošā citoplazma izskatās kā pavedieni, kas saplūst tikai ap kodolu. Gaisa kameras ir arī sifonoforiem, vairākām medūzām, planktona vēderkājiem u.c.
Jūras aļģes (fitoplanktons) pasīvi lidināties ūdenī, savukārt vairums planktona dzīvnieku spēj aktīvi peldēt, bet ierobežotās robežās. Planktona organismi nevar pārvarēt straumes, un tie tiek pārnesti lielos attālumos. Daudz veidu zooplanktons tomēr spēj veikt vertikālas migrācijas ūdens kolonnā desmitiem un simtiem metru gan aktīvas kustības, gan ķermeņa peldspējas regulēšanas dēļ. Īpašs planktona veids ir ekoloģiskā grupa Neuston ("Nein" - peldēt) - ūdens virsmas plēves iemītnieki pie robežas ar gaisa vidi.
Ūdens blīvums un viskozitāte lielā mērā ietekmē spēju aktīvi peldēt. Dzīvnieki, kas spēj ātri peldēt un pārvarēt straumju spēku, ir apvienoti ekoloģiskā grupā nekton ("Nektos" - peldošs). Nektona pārstāvji ir zivis, kalmāri, delfīni. Strauja kustība ūdens stabā iespējama tikai tad, ja ir racionālas ķermeņa formas un augsti attīstīti muskuļi. Torpēdas forma ir attīstīta visiem labiem peldētājiem, neatkarīgi no viņu sistemātiskās piederības un pārvietošanās metodes ūdenī: reaktīva, ķermeņa izliekuma dēļ, ar ekstremitāšu palīdzību.
Skābekļa režīms. Skābekļa ūdenī tā saturs nepārsniedz 10 ml uz 1 litru, kas ir 21 reizi mazāks nekā atmosfērā. Tāpēc ūdens organismu elpošanas apstākļi ir ievērojami sarežģīti. Skābeklis ūdenī nonāk galvenokārt aļģu fotosintēzes aktivitātes un difūzijas rezultātā no gaisa. Tāpēc ūdens staba augšējie slāņi, kā likums, ir bagātāki ar šo gāzi nekā apakšējie. Palielinoties temperatūrai un ūdens sāļumam, skābekļa koncentrācija tajā samazinās. Slāņos, kas ir ļoti apdzīvoti ar dzīvniekiem un baktērijām, tā palielinātā patēriņa dēļ var rasties straujš O 2 deficīts. Piemēram, Pasaules okeānā dzīvībai bagātos dziļumos no 50 līdz 1000 m raksturīga strauja aerācijas pasliktināšanās - tā ir 7-10 reizes zemāka nekā virszemes ūdeņos, ko apdzīvo fitoplanktons. Ūdenstilpju dibena tuvumā apstākļi var būt tuvu anaerobiem.
Starp ūdens iemītniekiem ir daudzas sugas, kas spēj panest lielas skābekļa satura svārstības ūdenī līdz pat gandrīz pilnīgai neesamībai. (eirooksibionti - "oksi" - skābeklis, "biont" - iedzīvotājs). Tajos ietilpst, piemēram, saldūdens oligošetes Tubifex tubifex un vēderkāji Viviparus viviparus. No zivīm ļoti vāju ūdens piesātinājumu ar skābekli var izturēt karpas, līņi un karūsas. Tomēr vairāki veidi stenoksibionti - tie var pastāvēt tikai ar pietiekami augstu ūdens piesātinājumu ar skābekli (varavīksnes forele, strauta forele, vīgrieze, skropstains tārps Planaria alpina, maijvaboļu kāpuri, akmeņmušiņas u.c.). Daudzas sugas spēj nonākt neaktīvā stāvoklī ar skābekļa trūkumu - anoksibioze - un tādējādi piedzīvo nelabvēlīgu periodu.
Ūdens organismu elpošana tiek veikta vai nu caur ķermeņa virsmu, vai arī caur specializētiem orgāniem - žaunām, plaušām, traheju. Šajā gadījumā apvalks var kalpot kā papildu elpošanas orgāns. Piemēram, sārta zivs caur ādu patērē līdz pat 63% skābekļa. Ja gāzu apmaiņa notiek caur ķermeņa daļām, tad tās ir ļoti plānas. Elpošanu atvieglo arī palielinātais virsmas laukums. Tas tiek panākts sugu evolūcijas laikā, veidojot dažādus izaugumus, saplacinot, pagarinot un vispārēji samazinot ķermeņa izmēru. Dažas sugas ar skābekļa trūkumu aktīvi maina elpošanas virsmas izmēru. Tārpi Tubifex tubifex spēcīgi izstiepj ķermeni; hidras un anemoni - taustekļi; adatādaiņi - ambulakrālās kājas. Daudzi mazkustīgi un mazkustīgi dzīvnieki atjauno ūdeni ap tiem, vai nu radot virzītu tā strāvu, vai arī ar svārstībām kustībām, kas veicina ūdens sajaukšanos. Šim nolūkam gliemenes ir skropstas, kas izklāj mantijas dobuma sienas; vēžveidīgie - vēdera vai krūšu kurvja kāju darbs. Dēles, zvana odu (asinstārpu) kāpuri, daudzas oligošetas šūpo ķermeni, izceļoties no zemes.
Dažām sugām tiek konstatēta ūdens un gaisa elpošanas kombinācija. Tās ir plaušas elpojošas zivis, sifonofori diskofanti, daudzi plaušu mīkstmieši, vēžveidīgie Gammarus lacustris u.c. Sekundārie dzīvnieki parasti saglabā atmosfēras elpošanas veidu kā enerģētiski izdevīgāku, un tāpēc tiem ir nepieciešami kontakti ar gaisa vidi, piemēram, roņveidīgajiem, vaļveidīgajiem, ūdens. vaboles, moskītu kāpuri un citi.
Skābekļa trūkums ūdenī dažkārt izraisa katastrofālas parādības - Zamora, ko pavada daudzu ūdens organismu nāve. Ziemas zamory bieži izraisa ledus veidošanās uz rezervuāru virsmas un kontakta ar gaisu pārtraukšana; vasara- ūdens temperatūras paaugstināšanās un skābekļa šķīdības samazināšanās rezultātā.
Bieža zivju un daudzu bezmugurkaulnieku bojāeja ziemā ir raksturīga, piemēram, Ob upes baseina lejasdaļai, kuras ūdeņi, kas plūst lejup no Rietumsibīrijas zemienes purvainajiem apgabaliem, ir ārkārtīgi nabadzīgi ar izšķīdušo skābekli. Dažreiz nāves gadījumi notiek jūrās.
Papildus skābekļa trūkumam nogalināšanu var izraisīt toksisko gāzu koncentrācijas palielināšanās ūdenī - metāns, sērūdeņradis, CO 2 u.c., kas veidojas organisko vielu sadalīšanās rezultātā ūdens apakšā. rezervuāri.
Sāls režīms.Ūdens organismu ūdens bilances uzturēšanai ir sava specifika. Ja sauszemes dzīvniekiem un augiem vissvarīgākais ir nodrošināt organismu ar ūdeni tā trūkuma apstākļos, tad ūdens organismiem tikpat svarīgi ir uzturēt noteiktu ūdens daudzumu organismā ar tā pārpalikumu vidē. Pārmērīgs ūdens daudzums šūnās izraisa osmotiskā spiediena izmaiņas un svarīgāko dzīvībai svarīgo funkciju traucējumus.
Lielākā daļa ūdensdzīvnieku poikilosmotisks: osmotiskais spiediens viņu ķermenī ir atkarīgs no apkārtējā ūdens sāļuma. Tāpēc galvenais ūdens organismu sāls līdzsvara saglabāšanas veids ir izvairīties no biotopiem ar neatbilstošu sāļumu. Saldūdens formas jūrās nevar pastāvēt, jūras formas nevar paciest atsāļošanu. Ja ūdens sāļums ir pakļauts izmaiņām, dzīvnieki pārvietojas, meklējot labvēlīgu vidi. Piemēram, jūras virszemes slāņu atsāļošanas laikā pēc spēcīgām lietavām radiolāri, jūras vēžveidīgie Calanus un citi nolaižas 100 m dziļumā.Mugurkaulnieki, augstākie vēži, kukaiņi un to kāpuri, kas dzīvo ūdenī, pieder homeosmotisks sugas, saglabājot nemainīgu osmotisko spiedienu organismā neatkarīgi no sāļu koncentrācijas ūdenī.
Saldūdens sugās ķermeņa sulas ir hipertoniskas attiecībā pret apkārtējo ūdeni. Tiem draud pārmērīga laistīšana, ja netiek novērsta liekā ūdens iekļūšana organismā vai izvadīšana no tā. Vienšūņiem tas tiek panākts ar ekskrēcijas vakuolu darbu, daudzšūnu organismos - izvadot ūdeni caur ekskrēcijas sistēmu. Daži ciliāti ik pēc 2-2,5 minūtēm izdala ūdens daudzumu, kas ir vienāds ar ķermeņa tilpumu. Šūna tērē daudz enerģijas, lai "izsūknētu" lieko ūdeni. Palielinoties sāļumam, vakuolu darbs palēninās. Tātad Paramecium apavos ar ūdens sāļumu 2,5% o vakuola pulsē ar intervālu 9 s, pie 5% o - 18 s, pie 7,5% o - 25 s. Pie sāls koncentrācijas 17,5% o vakuola pārstāj darboties, jo pazūd osmotiskā spiediena atšķirība starp šūnu un vidi.
Ja ūdens ir hipertonisks attiecībā pret ūdens organismu ķermeņa šķidrumiem, tiem ir dehidratācijas risks osmotisko zudumu rezultātā. Aizsardzība pret dehidratāciju tiek panākta, palielinot sāļu koncentrāciju ūdens organismu organismā. Dehidratāciju novērš homoiosmotisko organismu - zīdītāju, zivju, augstāko vēžu, ūdens kukaiņu un to kāpuru ūdensnecaurlaidīgie vāki.
Daudzas poikilosmotiskās sugas nonāk neaktīvā stāvoklī - apturēta animācija, jo organismā trūkst ūdens un palielinās sāļums. Tas ir raksturīgi sugām, kas mīt jūras ūdens peļķēs un piekrastes zonā: rotifers, flagellates, ciliates, daži vēžveidīgie, Melnās jūras daudzslāņu Nereis divesicolor u.c. Sāls anabioze- līdzeklis, lai izdzīvotu nelabvēlīgos periodos mainīga ūdens sāļuma apstākļos.
Patiesi eirihalīns starp ūdens iemītniekiem nav tik daudz sugu, kas aktīvā stāvoklī varētu dzīvot gan saldūdenī, gan sālsūdenī. Tās galvenokārt ir sugas, kas apdzīvo upju estuārus, estuārus un citas iesāļas ūdenstilpes.
Temperatūras režīmsūdenstilpes ir stabilākas nekā uz sauszemes. Tas ir saistīts ar ūdens fizikālajām īpašībām, pirmkārt, lielo īpatnējo siltumietilpību, kuras dēļ ievērojama siltuma daudzuma saņemšana vai izdalīšana neizraisa pārāk asas temperatūras izmaiņas. Ūdens iztvaikošana no rezervuāru virsmas, kurā tiek iztērēti aptuveni 2263,8 J/g, novērš apakšējo slāņu pārkaršanu, savukārt ledus veidošanās, kurā izdalās saplūšanas siltums (333,48 J/g), palēnina to darbību. dzesēšana.
Gada temperatūras svārstību amplitūda okeāna augšējos slāņos ir ne vairāk kā 10-15 ° C, kontinentālajās ūdenstilpēs - 30-35 ° C. Dziļūdens slāņiem ir raksturīga nemainīga temperatūra. Ekvatoriālajos ūdeņos virszemes slāņu vidējā gada temperatūra ir + (26-27) ° C, polārajos ūdeņos - aptuveni 0 ° C un zemāka. Karstos sauszemes avotos ūdens temperatūra var pietuvoties +100 ° C, un zemūdens geizeros pie augsta spiediena okeāna dibenā tiek reģistrēta +380 ° C temperatūra.
Tādējādi ūdenstilpēs ir diezgan liela temperatūras apstākļu dažādība. Starp augšējiem ūdens slāņiem ar tajos izteiktām sezonālām temperatūras svārstībām un apakšējiem, kur termiskais režīms ir nemainīgs, atrodas temperatūras lēciena zona jeb termoklīns. Termoklīns ir izteiktāks siltajās jūrās, kur temperatūras starpība starp ārējiem un dziļajiem ūdeņiem ir spēcīgāka.
Sakarā ar stabilāku ūdens temperatūras režīmu ūdens organismu vidū, stenoterma ir izplatīta daudz lielākā mērā nekā sauszemes populācijā. Eiritermālās sugas sastopamas galvenokārt seklās kontinentālās ūdenstilpēs un augstu un mēreno platuma grādu jūru piekrastē, kur ir būtiskas ikdienas un sezonālās temperatūras svārstības.
Gaismas režīms.Ūdenī ir daudz mazāk gaismas nekā gaisā. Daļa no stariem, kas nokrīt uz rezervuāra virsmas, tiek atspoguļoti gaisā. Jo zemāka ir Saules pozīcija, jo spēcīgāks ir atspīdums, tāpēc diena zem ūdens ir īsāka nekā uz sauszemes. Piemēram, vasaras diena pie Madeiras salas 30 m dziļumā - 5 stundas, bet 40 m dziļumā tikai 15 minūtes. Strauja gaismas daudzuma samazināšanās līdz ar dziļumu ir saistīta ar tās absorbciju ūdenī. Stari ar dažādu viļņu garumu netiek absorbēti vienādi: sarkanie pazūd jau tuvu virsmai, savukārt zili zaļie iekļūst daudz dziļāk. Krēsla, kas padziļinās okeānā, vispirms ir zaļa, tad zila, zila un zili violeta, beidzot dodot vietu pastāvīgai tumsai. Attiecīgi zaļās, brūnās un sarkanās aļģes, kas specializējas gaismas uztveršanā ar dažādu viļņu garumu, aizstāj viena otru ar dziļumu.
Dzīvnieku krāsa mainās līdz ar dziļumu tādā pašā veidā. Visspilgtāk un daudzveidīgāk krāsojas piekrastes un sublitorālās zonas iemītnieki. Daudziem dziļi iesakņojušajiem organismiem, piemēram, alu organismiem, nav pigmentu. Krēslas zonā plaši izplatīts sarkans krāsojums, kas šajos dziļumos papildina zili violeto gaismu. Krāsai papildinošos starus ķermenis absorbē vispilnīgāk. Tas ļauj dzīvniekiem paslēpties no ienaidniekiem, jo to sarkanā krāsa zili violetajos staros vizuāli tiek uztverta kā melna. Sarkanais krāsojums ir raksturīgs tādiem krēslas zonas dzīvniekiem kā jūras asaris, sarkanie koraļļi, dažādi vēžveidīgie u.c.
Dažām sugām, kas dzīvo netālu no ūdenstilpju virsmas, acis ir sadalītas divās daļās ar atšķirīgu spēju lauzt starus. Viena acs puse redz gaisā, otra ūdenī. Šāds "četracains" ir raksturīgs vabolēm-vītnēm, Amerikas zivīm Anableps tetraphthalmus, vienai no tropu jūras suņu sugām Dialommus fuscus. Paisuma laikā šī zivs sēž padziļinājumos, pakļaujot daļu galvas no ūdens (skat. 26. att.).
Gaismas absorbcija ir spēcīgāka, jo mazāka ir ūdens caurspīdīgums, kas ir atkarīgs no tajā suspendēto daļiņu daudzuma.
Caurspīdīgumu raksturo ārkārtējs dziļums, kurā joprojām ir redzams īpaši nolaists balts disks ar aptuveni 20 cm diametru (Secchi disks). Caurspīdīgākie ūdeņi ir Sargasu jūrā: disks ir redzams līdz 66,5 m dziļumam Klusajā okeānā Secchi disks ir redzams līdz 59 m, Indijas okeānā - līdz 50, seklajā jūrā - uz augšu. līdz 5-15 m Upju caurredzamība ir vidēji 1-1, 5 m, bet dubļainākajās upēs, piemēram, Vidusāzijas Amudarja un Sirdarja, tikai daži centimetri. Tāpēc fotosintēzes zonas robeža dažādās ūdenstilpēs ir ļoti atšķirīga. Tīrākajos ūdeņos eifotisks zona vai fotosintēzes zona sniedzas ne vairāk kā 200 m dziļumā, krēslā vai disfotisks, zona aizņem līdz 1000-1500 m dziļumu un dziļāk, afotisks zonā, saules gaisma neiekļūst vispār.
Gaismas daudzums ūdenstilpņu augšējos slāņos ir ļoti atšķirīgs atkarībā no teritorijas platuma un gadalaika. Garās polārās naktis ļoti ierobežo fotosintēzei pieejamo laiku Arktikas un Antarktikas baseinos, un ledus sega apgrūtina gaismai ziemā piekļūt visām sasalstošajām ūdenstilpēm.
Okeāna tumšajos dziļumos organismi izmanto dzīvu būtņu izstaroto gaismu kā vizuālās informācijas avotu. Par dzīvā organisma mirdzumu sauc bioluminiscence. Gaismas sugas ir sastopamas gandrīz visās ūdensdzīvnieku klasēs, sākot no vienšūņiem līdz zivīm, kā arī starp baktērijām, zemākajiem augiem un sēnēm. Bioluminiscence acīmredzot parādījās daudzas reizes dažādās grupās dažādos evolūcijas posmos.
Bioluminiscences ķīmija tagad ir diezgan labi saprotama. Gaismas radīšanai izmantotās reakcijas ir dažādas. Bet visos gadījumos tā ir sarežģītu organisko savienojumu oksidēšana. (luciferīni) izmantojot proteīnu katalizatorus (luciferāze). Luciferīniem un luciferāzēm dažādos organismos ir atšķirīga struktūra. Reakcijas laikā ierosinātās luciferīna molekulas enerģijas pārpalikums tiek atbrīvots gaismas kvantu veidā. Dzīvi organismi izstaro gaismu impulsu veidā, parasti reaģējot uz ārējās vides stimuliem.
Luminiscencei var nebūt īpašas ekoloģiskas nozīmes sugas dzīvē, bet tā var būt šūnu dzīvībai svarīgās aktivitātes blakusprodukts, kā, piemēram, baktērijās vai zemākajos augos. Tas iegūst ekoloģisko nozīmi tikai dzīvniekiem ar pietiekami attīstītu nervu sistēmu un redzes orgāniem. Daudzām sugām luminiscences orgāni iegūst ļoti sarežģītu struktūru ar atstarotāju un lēcu sistēmu, kas pastiprina starojumu (40. att.). Vairākas zivis un galvkāji, kas nespēj radīt gaismu, izmanto simbiotiskas baktērijas, kas vairojas īpašos šo dzīvnieku orgānos.
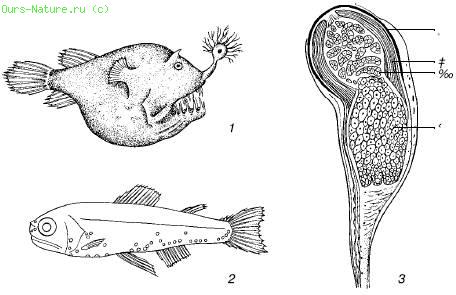
Rīsi. 40. Ūdensdzīvnieku mirdzošie orgāni (pēc S.A.Zernova, 1949):
1 - dziļjūras makšķernieks ar lukturīti virs zobainās mutes;
2 - gaismas orgānu izplatība zivīs ar šo. Mystophidae;
3 - zivju Argyropelecus affinis gaismas orgāns:
a - pigments, b - reflektors, c - gaismas korpuss, d - lēca
Bioluminiscencei galvenokārt ir signāla nozīme dzīvnieku dzīvē. Gaismas signāli var kalpot orientācijai barā, pievilinot pretējā dzimuma indivīdus, pievilinot upurus, maskējoties vai novēršot uzmanību. Gaismas zibspuldze var būt aizsardzība pret plēsēju, to apžilbinot vai dezorientējot. Piemēram, dziļjūras sēpijas, bēgot no ienaidnieka, izdala spoža sekrēta mākoni, savukārt apgaismotos ūdeņos dzīvojošās sugas šim nolūkam izmanto tumšu šķidrumu. Dažiem bentosa tārpiem - daudzspārņiem - gaismas orgāni attīstās līdz reproduktīvo produktu nobriešanas periodam, un tie spīd spožāk nekā mātīte, un vīriešiem ir labāk attīstītas acis. Plēsīgajām dziļjūras zivīm no jūrasvelnu kārtas pirmais muguras spuras stars tiek novirzīts uz augšžokli un pārvērsts par elastīgu "stieni", kura galā nes tārpveida "ēsmu" - dziedzeru, kas piepildīts ar gļotām ar gaismas baktērijas. Regulējot asins plūsmu uz dziedzeri un līdz ar to arī baktēriju piegādi ar skābekli, zivis var patvaļīgi izraisīt "ēsmas" spīdumu, imitējot tārpa kustības un pievilinot medījumu.