4. nodaļa. PAMATVIDES UN PIELĀGOŠANĀS ORGANISMI
4.1. Ūdens biotops. Ūdens organismu adaptācijas specifika
Ūdenim kā biotopam ir vairākas specifiskas īpašības, piemēram, augsts blīvums, spēcīgi spiediena kritumi, salīdzinoši zems skābekļa saturs, spēcīga saules gaismas absorbcija utt., suspendēto daļiņu saturs. Bentosa organismu dzīvībai svarīgas ir augsnes īpašības, organisko atlieku sadalīšanās veids uc Tāpēc līdz ar pielāgošanos vispārējām ūdens vides īpašībām tās iemītnieki ir jāpielāgo dažādiem īpašiem apstākļiem. Ūdens vides iedzīvotāji saņēma kopīgu nosaukumu ekoloģijā hidrobionti. Viņi apdzīvo okeānus, kontinentālās ūdenstilpes un gruntsūdeņus. Jebkurā ūdenstilpē ir iespējams izdalīt zonas, kas ir atšķirīgas apstākļu ziņā.
4.1.1. Pasaules okeāna ekoloģiskās zonas
Okeānā un tajā iekļautajās jūrās, pirmkārt, tiek izdalīti divi ekoloģiskie reģioni: ūdens stabs - pelagiāls un apakšā - bentāls (38. att.). Atkarībā no dziļuma bentāls tiek sadalīts sublitorāls zona - gludas zemes nolaišanās apgabals aptuveni 200 m dziļumā, batiāls- stāvas nogāzes laukums un bezdibeņa zona- okeāna gultnes laukums ar vidējo dziļumu 3–6 km. Tiek saukti pat dziļāki bentāla apgabali, kas atbilst okeāna gultnes ieplakas ultraabisāls. Paisuma un paisuma laikā applūdušo krasta malu sauc piekraste. Virs paisuma līmeņa sauc to piekrastes daļu, kas samitrināta ar sērfa aerosolu supralitorāls.
Rīsi. 38. Pasaules okeāna ekoloģiskās zonas
Dabiski, piemēram, sublitorāla iemītnieki dzīvo relatīvi zema spiediena, dienas saules gaismas un nereti visai ievērojamu temperatūras izmaiņu apstākļos. Bezdibeņa un īpaši bezdibeņa dzīļu iemītnieki pastāv tumsā, nemainīgā temperatūrā un milzīgā spiedienā, kas sasniedz vairākus simtus un dažreiz apmēram tūkstoš atmosfēru. Tāpēc jau norāde par to, kurā bentāla zonā dzīvo konkrēta organismu suga, jau liecina par to, kādām vispārējām ekoloģiskajām īpašībām tai vajadzētu būt. Tika nosaukti visi okeāna dibena iedzīvotāji bentoss.
Organismi, kas dzīvo ūdens kolonnā jeb pelagiāli, pieder pie pelagos. Pelagiāls ir sadalīts arī vertikālās zonās, kas dziļumā atbilst bentālajām zonām: epipelaģisks, batipelaģisks, abysopelagial. Epipelaģiskās zonas apakšējo robežu (ne vairāk kā 200 m) nosaka saules gaismas iekļūšana fotosintēzei pietiekamā daudzumā. Fotosintētiskie augi nevar pastāvēt dziļāk par šīm zonām. Krēslas batiālā un tumsas pilnajā bezdibeņa dziļumā dzīvo tikai mikroorganismi un dzīvnieki. Visos citos ūdenstilpņu veidos: ezeros, purvos, dīķos, upēs uc izšķir dažādas ekoloģiskās zonas. Visus šos biotopus apguvušo ūdens organismu daudzveidība ir ļoti liela.
4.1.2. Ūdens vides pamatīpašības
Ūdens blīvums- Tas ir faktors, kas nosaka ūdens organismu pārvietošanās apstākļus un spiedienu dažādos dziļumos. Destilētam ūdenim blīvums ir 1 g / cm 3 4 ° C temperatūrā. Dabisko ūdeņu, kas satur izšķīdušos sāļus, blīvums var būt lielāks, līdz 1,35 g / cm 3. Spiediens palielinās līdz ar dziļumu par aptuveni 1 · 10 5 Pa (1 atm) vidēji uz katriem 10 m.
Krasā spiediena gradienta dēļ ūdenstilpēs ūdens organismi parasti ir daudz eirobātiskāki nekā sauszemes organismi. Dažas sugas, kas izplatītas dažādos dziļumos, pieļauj spiedienu no vairākiem līdz simtiem atmosfēru. Piemēram, Elpidia ģints jūras gurķi, Priapulus caudatus tārpi apdzīvo no piekrastes zonas līdz ultraabyssal. Pat saldūdens iemītnieki, piemēram, skropstiņi, čības, suvoji, peldvaboles utt., eksperimentā var izturēt līdz 6 · 10 7 Pa (600 atm).
Tomēr daudzi jūru un okeānu iedzīvotāji ir relatīvi stenobāti un atrodas noteiktā dziļumā. Stenobātisms visbiežāk ir raksturīgs sekla un dziļūdens sugām. Tikai piekrastes joslu apdzīvo annelidsmilšu tārps Arenicola un jūras šķīvīši (Patella). Daudzas zivis, piemēram, no makšķernieku grupas, galvkāji, vēžveidīgie, pogonofori, jūras zvaigznes u.c. sastopamas tikai lielā dziļumā vismaz 4 · 10 7 - 5 · 10 7 Pa (400–500 atm) spiedienā.
Ūdens blīvums nodrošina iespēju uz to paļauties, kas ir īpaši svarīgi skeleta formām. Vides blīvums kalpo kā nosacījums planēšanai ūdenī, un daudzi ūdens organismi ir pielāgoti šim konkrētajam dzīvesveidam. Suspendētie, ūdenī peldošie organismi tiek apvienoti īpašā ekoloģiskā ūdens organismu grupā - planktons ("Planktos" - planējošs).
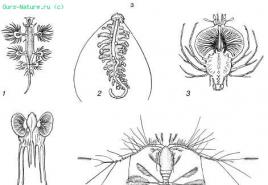
Rīsi. 39. Ķermeņa relatīvās virsmas palielināšanās planktona organismos (saskaņā ar S. A. Zernovu, 1949):
A — stieņa formas:
1 - diatoma Synedra;
2 - zilaļģes Aphanizomenon;
3 - Peridineal aļģes Amphisolenia;
4 - Euglena acus;
5 - galvkāji Doratopsis vermicularis;
6 - copepod Setella;
7 - Porcelānas kāpurs (Decapoda)
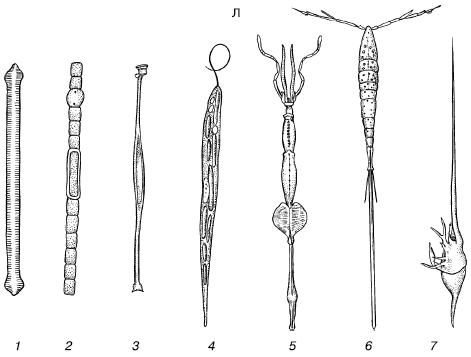
B — sadalītas formas:
1 - gliemji Glaucus atlanticus;
2 - tārps Tomopetris euchaeta;
3 - vēžu Palinurus kāpurs;
4 - jūrasvelna Lophius zivju kāpurs;
5 - copepod Calocalanus pavo
Planktonā ir vienšūnu un koloniālās aļģes, vienšūņi, medūzas, sifonofori, ķemmes želejas, pterigopodi un gliemenes, dažādi mazie vēžveidīgie, bentosa dzīvnieku kāpuri, zivju ikri un mazuļi un daudzi citi (39. att.). Planktona organismiem ir daudz līdzīgu pielāgojumu, kas palielina to peldspēju un neļauj tiem nogrimt dibenā. Šādi pielāgojumi ietver: 1) vispārēju ķermeņa relatīvās virsmas palielināšanos izmēru samazināšanās, saplacināšanas, pagarinājuma, daudzu izaugumu vai saru veidošanās dēļ, kas palielina berzi pret ūdeni; 2) blīvuma samazināšanās, ko izraisa skeleta samazināšanās, tauku uzkrāšanās, gāzu burbuļi organismā utt. Kramaļmatos rezerves vielas nogulsnējas nevis smagās cietes, bet gan tauku pilienu veidā. Nightlight Noctiluca izceļas ar tādu gāzu vakuolu un tauku pilienu pārpilnību šūnā, ka tajā esošā citoplazma izskatās kā pavedieni, kas saplūst tikai ap kodolu. Gaisa kameras ir arī sifonoforiem, vairākām medūzām, planktona vēderkājiem u.c.
Jūras aļģes (fitoplanktons) pasīvi lidināties ūdenī, savukārt vairums planktona dzīvnieku spēj aktīvi peldēt, bet ierobežotās robežās. Planktona organismi nevar pārvarēt straumes, un tie tiek pārnesti lielos attālumos. Daudz veidu zooplanktons tomēr spēj veikt vertikālas migrācijas ūdens kolonnā desmitiem un simtiem metru gan aktīvas kustības, gan ķermeņa peldspējas regulēšanas dēļ. Īpašs planktona veids ir ekoloģiskā grupa Neuston ("Nein" - peldēt) - ūdens virsmas plēves iemītnieki pie robežas ar gaisa vidi.
Ūdens blīvums un viskozitāte lielā mērā ietekmē spēju aktīvi peldēt. Dzīvnieki, kas spēj ātri peldēt un pārvarēt straumju spēku, ir apvienoti ekoloģiskā grupā nekton ("Nektos" - peldošs). Nektona pārstāvji ir zivis, kalmāri, delfīni. Strauja kustība ūdens stabā iespējama tikai tad, ja ir racionālas ķermeņa formas un augsti attīstīti muskuļi. Torpēdas forma ir attīstīta visiem labiem peldētājiem, neatkarīgi no viņu sistemātiskās piederības un pārvietošanās metodes ūdenī: reaktīva, ķermeņa izliekuma dēļ, ar ekstremitāšu palīdzību.
Skābekļa režīms. Skābekļa ūdenī tā saturs nepārsniedz 10 ml uz 1 litru, kas ir 21 reizi mazāks nekā atmosfērā. Tāpēc ūdens organismu elpošanas apstākļi ir ievērojami sarežģīti. Skābeklis ūdenī nonāk galvenokārt aļģu fotosintēzes aktivitātes un difūzijas rezultātā no gaisa. Tāpēc ūdens staba augšējie slāņi, kā likums, ir bagātāki ar šo gāzi nekā apakšējie. Palielinoties temperatūrai un ūdens sāļumam, skābekļa koncentrācija tajā samazinās. Slāņos, kas ir ļoti apdzīvoti ar dzīvniekiem un baktērijām, tā pieaugošā patēriņa dēļ var rasties straujš O 2 deficīts. Piemēram, Pasaules okeānā dzīvībai bagātos dziļumos no 50 līdz 1000 m raksturīga strauja aerācijas pasliktināšanās - tā ir 7-10 reizes zemāka nekā virszemes ūdeņos, ko apdzīvo fitoplanktons. Ūdenstilpju dibena tuvumā apstākļi var būt tuvu anaerobiem.
Starp ūdens iemītniekiem ir daudzas sugas, kas spēj panest lielas skābekļa satura svārstības ūdenī līdz pat gandrīz pilnīgai neesamībai. (eirooksibionti - "oksi" - skābeklis, "biont" - iedzīvotājs). Tajos ietilpst, piemēram, saldūdens oligošetes Tubifex tubifex un vēderkāji Viviparus viviparus. No zivīm ļoti vāju ūdens piesātinājumu ar skābekli var izturēt karpas, līņi un karūsas. Tomēr vairāki veidi stenoksibionti - tie var pastāvēt tikai ar pietiekami augstu ūdens piesātinājumu ar skābekli (varavīksnes forele, strauta forele, vīgrieze, skropstains tārps Planaria alpina, maijvaboļu kāpuri, akmeņmušiņas u.c.). Daudzas sugas spēj nonākt neaktīvā stāvoklī ar skābekļa trūkumu - anoksibioze - un tādējādi piedzīvo nelabvēlīgu periodu.
Ūdens organismu elpošana tiek veikta vai nu caur ķermeņa virsmu, vai caur specializētiem orgāniem - žaunām, plaušām, traheju. Šajā gadījumā apvalks var kalpot kā papildu elpošanas orgāns. Piemēram, sārta zivs caur ādu patērē līdz pat 63% skābekļa. Ja gāzu apmaiņa notiek caur ķermeņa daļām, tad tās ir ļoti plānas. Elpošanu atvieglo arī palielinātais virsmas laukums. Tas tiek panākts sugu evolūcijas laikā, veidojot dažādus izaugumus, saplacinot, pagarinot un vispārēji samazinot ķermeņa izmēru. Dažas sugas ar skābekļa trūkumu aktīvi maina elpošanas virsmas izmēru. Tārpi Tubifex tubifex spēcīgi izstiepj ķermeni; hidras un anemoni - taustekļi; adatādaiņi - ambulakrālās kājas. Daudzi mazkustīgi un mazkustīgi dzīvnieki atjauno ūdeni ap tiem, vai nu radot virzītu tā strāvu, vai arī ar svārstībām kustībām, kas veicina ūdens sajaukšanos. Šim nolūkam gliemenes ir skropstas, kas izklāj mantijas dobuma sienas; vēžveidīgie - vēdera vai krūšu kurvja kāju darbs. Dēles, zvana odu (asinstārpu) kāpuri, daudzas oligohetas šūpo ķermeni, noliecoties no zemes.
Dažām sugām tiek konstatēta ūdens un gaisa elpošanas kombinācija. Tās ir plaušas elpojošas zivis, sifonofori diskofanti, daudzi plaušu mīkstmieši, vēžveidīgie Gammarus lacustris u.c. Sekundārie dzīvnieki parasti saglabā atmosfēras elpošanas veidu kā enerģētiski izdevīgāku, tāpēc tiem ir nepieciešami kontakti ar gaisa vidi, piemēram, roņveidīgajiem, vaļveidīgajiem, ūdens. vaboles, moskītu kāpuri un citi.
Skābekļa trūkums ūdenī dažkārt izraisa katastrofālas parādības - Zamora, ko pavada daudzu ūdens organismu nāve. Ziemas zamory bieži izraisa ledus veidošanās uz rezervuāru virsmas un kontakta ar gaisu pārtraukšana; vasara- ūdens temperatūras paaugstināšanās un skābekļa šķīdības samazināšanās rezultātā.
Bieža zivju un daudzu bezmugurkaulnieku bojāeja ziemā ir raksturīga, piemēram, Ob upes baseina lejasdaļai, kuras ūdeņi, kas plūst lejup no Rietumsibīrijas zemienes purvainajiem apgabaliem, ir ārkārtīgi nabadzīgi ar izšķīdušo skābekli. Dažreiz nāves gadījumi notiek jūrās.
Papildus skābekļa trūkumam salnas var izraisīt toksisko gāzu koncentrācijas palielināšanās ūdenī - metāns, sērūdeņradis, CO 2 u.c., kas veidojas organisko vielu sadalīšanās rezultātā ūdens apakšā. rezervuāri.
Sāls režīms.Ūdens organismu ūdens bilances uzturēšanai ir sava specifika. Ja sauszemes dzīvniekiem un augiem vissvarīgākais ir nodrošināt organismu ar ūdeni tā trūkuma apstākļos, tad ūdens organismiem tikpat svarīgi ir uzturēt noteiktu ūdens daudzumu organismā ar tā pārpalikumu vidē. Pārmērīgs ūdens daudzums šūnās izraisa osmotiskā spiediena izmaiņas un svarīgāko dzīvībai svarīgo funkciju traucējumus.
Lielākā daļa ūdensdzīvnieku poikilosmotisks: osmotiskais spiediens viņu ķermenī ir atkarīgs no apkārtējā ūdens sāļuma. Tāpēc galvenais ūdens organismu sāls līdzsvara saglabāšanas veids ir izvairīties no biotopiem ar neatbilstošu sāļumu. Saldūdens formas jūrās nevar pastāvēt, jūras formas nevar paciest atsāļošanu. Ja ūdens sāļums ir pakļauts izmaiņām, dzīvnieki pārvietojas, meklējot labvēlīgu vidi. Piemēram, jūras virszemes slāņu atsāļošanas laikā pēc stiprām lietavām 100 m dziļumā nolaižas radiolāri, jūras vēžveidīgie Calanus u.c.. Mugurkaulnieki, augstākie vēži, kukaiņi un to kāpuri, kas dzīvo ūdenī, pieder homeosmotisks sugas, saglabājot nemainīgu osmotisko spiedienu organismā neatkarīgi no sāļu koncentrācijas ūdenī.
Saldūdens sugās ķermeņa sulas ir hipertoniskas attiecībā pret apkārtējo ūdeni. Tiem draud pārmērīga laistīšana, ja netiek novērsta liekā ūdens iekļūšana organismā vai izvadīšana no tā. Vienšūņiem tas tiek panākts ar ekskrēcijas vakuolu darbu, daudzšūnu organismos - izvadot ūdeni caur ekskrēcijas sistēmu. Daži ciliāti ik pēc 2–2,5 minūtēm izdala ūdens daudzumu, kas vienāds ar ķermeņa tilpumu. Šūna tērē daudz enerģijas, lai "izsūknētu" lieko ūdeni. Palielinoties sāļumam, vakuolu darbs palēninās. Tātad Paramecium apavos ar ūdens sāļumu 2,5% o vakuola pulsē ar intervālu 9 s, pie 5% o - 18 s, pie 7,5% o - 25 s. Pie sāls koncentrācijas 17,5% o vakuola pārstāj darboties, jo pazūd osmotiskā spiediena atšķirība starp šūnu un vidi.
Ja ūdens ir hipertonisks attiecībā pret ūdens organismu ķermeņa šķidrumiem, tiem ir dehidratācijas risks osmotisko zudumu rezultātā. Aizsardzība pret dehidratāciju tiek panākta, palielinot sāļu koncentrāciju ūdens organismu organismā. Dehidratāciju novērš ūdensnecaurlaidīgi homoiosmotisko organismu vāki - zīdītāji, zivis, augstākie vēži, ūdens kukaiņi un to kāpuri.
Daudzas poikilosmotiskās sugas nonāk neaktīvā stāvoklī - apturēta animācija, jo organismā trūkst ūdens un palielinās sāļums. Tas ir raksturīgs sugām, kas mīt jūras ūdens peļķēs un piekrastes zonā: rotifers, flagellates, ciliates, daži vēžveidīgie, Melnās jūras daudzslāņu Nereis divesicolor u.c. Sāls anabioze- līdzeklis, lai izdzīvotu nelabvēlīgos periodos mainīga ūdens sāļuma apstākļos.
Patiesi eirihalīns starp ūdens iemītniekiem nav tik daudz sugu, kas aktīvā stāvoklī varētu dzīvot gan saldūdenī, gan sālsūdenī. Tās galvenokārt ir sugas, kas apdzīvo upju estuārus, estuārus un citas iesāļas ūdenstilpes.
Temperatūras režīmsūdenstilpes ir stabilākas nekā uz sauszemes. Tas ir saistīts ar ūdens fizikālajām īpašībām, pirmkārt, lielo īpatnējo siltumietilpību, kuras dēļ ievērojama siltuma daudzuma saņemšana vai izdalīšana neizraisa pārāk asas temperatūras izmaiņas. Ūdens iztvaikošana no rezervuāru virsmas, kurā tiek iztērēti aptuveni 2263,8 J/g, novērš apakšējo slāņu pārkaršanu, savukārt ledus veidošanās, kurā izdalās saplūšanas siltums (333,48 J/g), palēnina to darbību. dzesēšana.
Gada temperatūras svārstību amplitūda okeāna augšējos slāņos ir ne vairāk kā 10–15 ° C, kontinentālajās ūdenstilpēs - 30–35 ° C. Dziļūdens slāņiem ir raksturīga nemainīga temperatūra. Ekvatoriālajos ūdeņos virszemes slāņu vidējā gada temperatūra ir + (26–27) ° С, polārajos ūdeņos - aptuveni 0 ° C un zemāka. Karstos sauszemes avotos ūdens temperatūra var pietuvoties +100 ° C, un zemūdens geizeros pie augsta spiediena okeāna dibenā tiek reģistrēta +380 ° C temperatūra.
Tādējādi ūdenstilpēs ir diezgan liela temperatūras apstākļu dažādība. Starp augšējiem ūdens slāņiem ar tajos izteiktām sezonālām temperatūras svārstībām un apakšējiem, kur termiskais režīms ir nemainīgs, ir temperatūras lēciena zona jeb termoklīns. Termoklīns ir izteiktāks siltajās jūrās, kur temperatūras starpība starp ārējiem un dziļajiem ūdeņiem ir spēcīgāka.
Sakarā ar stabilāku ūdens temperatūras režīmu ūdens organismu vidū, stenoterma ir izplatīta daudz lielākā mērā nekā sauszemes populācijā. Eiritermālās sugas galvenokārt sastopamas seklās kontinentālās ūdenstilpēs un augstos un mērenos platuma grādos jūru piekrastē, kur ir būtiskas ikdienas un sezonālās temperatūras svārstības.
Gaismas režīms.Ūdenī ir daudz mazāk gaismas nekā gaisā. Daļa no stariem, kas nokrīt uz rezervuāra virsmas, tiek atspoguļoti gaisā. Jo zemāka ir Saules pozīcija, jo spēcīgāks ir atspīdums, tāpēc diena zem ūdens ir īsāka nekā uz sauszemes. Piemēram, vasaras diena pie Madeiras salas 30 m dziļumā - 5 stundas, bet 40 m dziļumā tikai 15 minūtes. Strauja gaismas daudzuma samazināšanās līdz ar dziļumu ir saistīta ar tās absorbciju ūdenī. Stari ar dažādu viļņu garumu tiek absorbēti atšķirīgi: sarkanie pazūd jau tuvu virsmai, savukārt zili zaļie iekļūst daudz dziļāk. Krēsla, kas padziļinās ar dziļumu okeānā, vispirms ir zaļa, tad zila, zila un zili violeta, beidzot dodot vietu pastāvīgai tumsai. Attiecīgi zaļās, brūnās un sarkanās aļģes, kas specializējas gaismas uztveršanā ar dažādu viļņu garumu, aizstāj viena otru ar dziļumu.
Dzīvnieku krāsa mainās līdz ar dziļumu tādā pašā veidā. Visspilgtāk un daudzveidīgāk krāsojas piekrastes un sublitorālās zonas iemītnieki. Daudziem dziļi iesakņojušajiem organismiem, piemēram, alu organismiem, nav pigmentu. Krēslas zonā plaši izplatīts sarkans krāsojums, kas šajos dziļumos papildina zili violeto gaismu. Krāsai papildinošos starus ķermenis absorbē vispilnīgāk. Tas ļauj dzīvniekiem paslēpties no ienaidniekiem, jo to sarkanā krāsa zili violetajos staros vizuāli tiek uztverta kā melna. Sarkanais krāsojums ir raksturīgs tādiem krēslas zonas dzīvniekiem kā jūras asaris, sarkanie koraļļi, dažādi vēžveidīgie u.c.
Dažām sugām, kas dzīvo netālu no ūdenstilpju virsmas, acis ir sadalītas divās daļās ar atšķirīgu spēju lauzt starus. Viena acs puse redz gaisā, otra ūdenī. Šāds "četracs" ir raksturīgs virpuļvabolēm, Amerikas zivīm Anableps tetraphthalmus, vienai no tropu jūras suņu Dialommus fuscus sugām. Paisuma laikā šī zivs sēž padziļinājumos, pakļaujot daļu galvas no ūdens (skat. 26. att.).
Gaismas absorbcija ir spēcīgāka, jo mazāka ir ūdens caurspīdīgums, kas ir atkarīgs no tajā suspendēto daļiņu daudzuma.
Caurspīdīgumu raksturo ārkārtējs dziļums, kurā joprojām ir redzams īpaši nolaists balts disks ar aptuveni 20 cm diametru (Secchi disks). Caurspīdīgākie ūdeņi ir Sargasu jūrā: disks ir redzams līdz 66,5 m dziļumam Klusajā okeānā Secchi disks ir redzams līdz 59 m, Indijas okeānā - līdz 50, seklajā jūrā - uz augšu. līdz 5-15 m.. Upju caurspīdība vidēji ir 1-1,5 m, bet dubļainākajās upēs, piemēram, Vidusāzijas Amudarja un Sirdarja, tikai daži centimetri. Tāpēc fotosintēzes zonas robeža dažādās ūdenstilpēs ir ļoti atšķirīga. Tīrākajos ūdeņos eifotisks zona vai fotosintēzes zona sniedzas ne vairāk kā 200 m dziļumā, krēslā vai disfotisks, zona aizņem līdz 1000-1500 m dziļumu un dziļāk, afotisks zonā, saules gaisma neiekļūst vispār.
Gaismas daudzums ūdenstilpņu augšējos slāņos ir ļoti atšķirīgs atkarībā no teritorijas platuma un gadalaika. Garās polārās naktis ļoti ierobežo fotosintēzei pieejamo laiku Arktikas un Antarktikas baseinos, un ledus sega apgrūtina gaismai ziemā piekļūt visām sasalstošajām ūdenstilpēm.
Okeāna tumšajos dziļumos organismi izmanto dzīvu būtņu izstaroto gaismu kā vizuālās informācijas avotu. Par dzīvā organisma mirdzumu sauc bioluminiscence. Gaismas sugas ir sastopamas gandrīz visās ūdensdzīvnieku klasēs, sākot no vienšūņiem līdz zivīm, kā arī starp baktērijām, zemākajiem augiem un sēnēm. Šķiet, ka bioluminiscence ir radusies daudzas reizes dažādās grupās dažādos evolūcijas posmos.
Bioluminiscences ķīmija tagad ir diezgan labi saprotama. Gaismas radīšanai izmantotās reakcijas ir dažādas. Bet visos gadījumos tā ir sarežģītu organisko savienojumu oksidēšana. (luciferīni) izmantojot proteīnu katalizatorus (luciferāze). Luciferīniem un luciferāzēm dažādos organismos ir atšķirīga struktūra. Reakcijas laikā ierosinātās luciferīna molekulas enerģijas pārpalikums tiek atbrīvots gaismas kvantu veidā. Dzīvi organismi izstaro gaismu impulsu veidā, parasti reaģējot uz ārējās vides stimuliem.
Luminiscencei var nebūt īpašas ekoloģiskas nozīmes sugas dzīvē, bet tā ir šūnu dzīvībai svarīgās aktivitātes blakusprodukts, kā, piemēram, baktērijās vai zemākajos augos. Tas iegūst ekoloģisko nozīmi tikai dzīvniekiem ar pietiekami attīstītu nervu sistēmu un redzes orgāniem. Daudzām sugām luminiscences orgāni iegūst ļoti sarežģītu struktūru ar atstarotāju un lēcu sistēmu, kas pastiprina starojumu (40. att.). Vairākas zivis un galvkāji, kas nespēj radīt gaismu, izmanto simbiotiskas baktērijas, kas vairojas īpašos šo dzīvnieku orgānos.
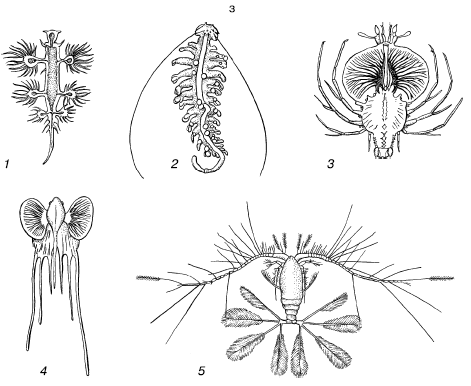
Rīsi. 40. Ūdensdzīvnieku mirdzošie orgāni (pēc S.A.Zernova, 1949):
1 - dziļjūras makšķernieks ar lukturīti virs zobainās mutes;
2 - gaismas orgānu izplatība zivīs. Mystophidae;
3 - zivju Argyropelecus affinis gaismas orgāns:
a - pigments, b - reflektors, c - gaismas korpuss, d - lēca
Bioluminiscencei galvenokārt ir signāla nozīme dzīvnieku dzīvē. Gaismas signāli var kalpot orientācijai barā, pievilinot pretējā dzimuma indivīdus, pievilinot upurus, maskējoties vai novēršot uzmanību. Gaismas zibspuldze var būt aizsardzība pret plēsēju, to apžilbinot vai dezorientējot. Piemēram, dziļjūras sēpijas, bēgot no ienaidnieka, izdala gaismas sekrēta mākoni, savukārt sugas, kas dzīvo apgaismotos ūdeņos, šim nolūkam izmanto tumšu šķidrumu. Dažiem bentosa tārpiem - daudzspārņiem - gaismas orgāni attīstās līdz reproduktīvo produktu nobriešanas periodam, un tie spīd spožāk nekā mātīte, un acis ir labāk attīstītas vīriešiem. Plēsīgajām dziļjūras zivīm no jūrasvelnu kārtas pirmais muguras spuras stars tiek novirzīts uz augšžokli un pārvērsts par elastīgu "stieni", kura galā nes tārpveida "ēsmu" - dziedzeru, kas piepildīts ar gļotām ar gaismas baktērijas. Regulējot asins plūsmu uz dziedzera un līdz ar to arī skābekļa piegādi baktērijām, zivs var patvaļīgi izraisīt “ēsmas” spīdumu, imitējot tārpa kustības un pievilinot medījumu.
Sauszemes vidē bioluminiscence attīstās tikai dažām sugām, visvairāk ugunspuķu dzimtas vabolēm, kuras ar gaismas signālu palīdzību pievilina pretējā dzimuma īpatņus krēslā vai naktī.
4.1.3. Daži specifiski ūdens organismu pielāgojumi
Metodes dzīvnieku orientēšanai ūdens vidē. Dzīvošana pastāvīgā krēslā vai tumsā stipri ierobežo iespējas vizuālā orientācija hidrobionti. Sakarā ar straujo gaismas staru vājināšanos ūdenī, pat labi attīstītu redzes orgānu īpašnieki tiek vadīti ar viņu palīdzību tikai no tuva attāluma.
Skaņa ūdenī izplatās ātrāk nekā gaisā. Skaņas orientācija parasti ir labāk attīstīts ūdens organismos nekā vizuālais. Vairāki veidi pat uztver ļoti zemas frekvences vibrācijas (infraskaņas), rodas, mainoties viļņu ritmam, un pirms vētras nolaižas no virszemes slāņiem uz dziļākiem (piemēram, medūzām). Daudzi ūdenstilpņu iemītnieki – zīdītāji, zivis, mīkstmieši, vēžveidīgie – paši izdod skaņas. Vēžveidīgie to dara, berzējot dažādas ķermeņa daļas vienu pret otru; zivis - ar peldpūšļa, rīkles zobu, žokļu, krūšu spuru staru palīdzību un citos veidos. Skaņas signalizācija visbiežāk tiek izmantota starpsugu attiecībām, piemēram, orientēšanās barā, pretējā dzimuma indivīdu piesaiste u.tml., un īpaši attīstīta duļķainu ūdeņu un lielu dziļumu iemītniekiem, kas dzīvo tumsā.
Vairāki ūdens organismi atrod barību un pārvietojas, izmantojot eholokācija- atstaroto skaņas viļņu uztvere (vaļveidīgie). Daudzi uztvert atstarotos elektriskos impulsus, radot dažādas frekvences izlādes peldēšanas laikā. Ir zināmas aptuveni 300 zivju sugas, kas ražo elektrību un izmanto to orientācijai un signalizācijai. Saldūdens ziloņu zivs (Mormyrus kannume) raida līdz 30 impulsiem sekundē, atklājot bezmugurkaulniekus, kurus tā medī šķidrās dūņās bez redzes palīdzības. Dažām jūras zivīm izplūdes biežums sasniedz 2000 impulsus sekundē. Vairākas zivis izmanto arī elektriskos laukus aizsardzībai un uzbrukumam (elektriskais stars, elektriskais zutis utt.).
Orientēšanās padziļinātai lietošanai hidrostatiskā spiediena uztvere. To veic ar statocistu, gāzes kameru un citu orgānu palīdzību.
Senākais orientēšanās veids, kas raksturīgs visiem ūdens dzīvniekiem, ir vides ķīmijas uztvere. Daudzu ūdens organismu ķīmiskie receptori ir ārkārtīgi jutīgi. Daudzām zivju sugām raksturīgajās tūkstoš kilometru migrācijās tās vadās galvenokārt pēc smakām, ar pārsteidzošu precizitāti atrodot nārsta vai barošanās vietas. Eksperimentāli ir pierādīts, piemēram, ka laši, kuriem mākslīgi atņemta oža, neatrod savas upes grīvu, atgriežoties nārstot, taču viņi nekad nekļūdās, ja spēj uztvert smakas. Smaržas smalkums ir īpaši izteikts zivīm, kas veic īpaši tālu migrāciju.
Pielāgošanās dzīvei izžūšanas ūdenstilpēs specifika. Uz Zemes ir daudz īslaicīgu, seklu ūdenstilpņu, kas rodas pēc upju applūšanas, stiprām lietavām, sniega kušanas uc Šajās ūdenstilpēs, neskatoties uz to eksistences īsumu, apmetas dažādi ūdens organismi.
Izžūstošo baseinu iemītnieku kopīgās iezīmes ir spēja īsā laikā dot daudz pēcnācēju un ilgstoši izturēt bez ūdens. Tajā pašā laikā daudzu sugu pārstāvji tiek aprakti dūņās, nonākot samazinātas dzīvības aktivitātes stāvoklī - hipobioze.Šādi rīkojas shtitti, kladocerāni, planārija, sīksaru tārpi, mīkstmieši un pat zivis - lops, Āfrikas protopterus un Dienvidamerikas lepidosirēns no plaušām. Daudzas mazas sugas veido cistas, kas var izturēt sausumu, piemēram, saulespuķes, ciliāti, sakneņi, virkne copepods, turbellaria un Rhabditis ģints nematodes. Citi piedzīvo nelabvēlīgu periodu ļoti izturīgu olu stadijā. Visbeidzot, dažiem mazajiem izžūstošo ūdenstilpņu iemītniekiem ir unikāla spēja izžūt līdz plēves stāvoklim un, samitrinot, atsāk augšanu un attīstību. Spēja izturēt pilnīgu ķermeņa dehidratāciju atklāja Callidina, Philodina uc ģints rotiferiem, Macrobiotus, Echiniscus tardigrades, Tylenchus, Plectus, Cephalobus ģints nematodēm uc Šie dzīvnieki apdzīvo mikrorezervuārus sūnām un ķērpjiem un ir pielāgoti pēkšņām mitruma režīma izmaiņām.
Filtrēšana kā pārtikas veids. Daudziem ūdens organismiem ir īpašs barošanās veids - tas ir ūdenī suspendētu organiskas izcelsmes daļiņu un daudzu mazu organismu celms vai sedimentācija (41. att.).

Rīsi. 41. Barenca jūras ascīdiešu planktona barības sastāvs (pēc S.A.Zernova, 1949)
Šāds barošanas veids, kas neprasa lielus enerģijas izdevumus medījuma meklējumos, ir raksturīgs slāņainajiem žaunu mīkstmiešiem, sēdošajiem adatādaiņiem, daudzveidīgajiem, bryozoiem, ascīdiem, planktona vēžveidīgajiem u.c. (42. att.). Dzīvniekiem, kas barojas ar filtru, ir būtiska nozīme ūdenstilpju bioloģiskajā attīrīšanā. Gliemenes, kas dzīvo 1 m 2 platībā, diennaktī caur mantijas dobumu var izvadīt 150–280 m 3 ūdens, izgulsnējot suspendētās daļiņas. Saldūdens dafnijas, ciklopi vai visizplatītākais vēžveidīgais Calanus finmarchicus okeānā filtrē līdz 1,5 litriem ūdens uz vienu indivīdu dienā. Okeāna piekrastes zona, kas ir īpaši bagāta ar filtrējošo organismu uzkrājumiem, darbojas kā efektīva attīrīšanas sistēma.
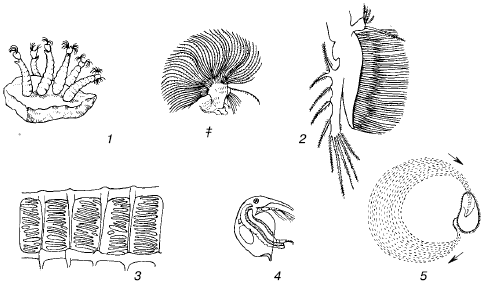
Rīsi. 42. Ūdens organismu filtrēšanas aparāts (pēc S.A.Zernova, 1949):
1 - Simulium midges kāpuri uz akmens (a) un to filtrējošie piedēkļi (b);
2 - vēžveidīgo Diaphanosoma brachyurum filtrējošā kāja;
3 - ascidian Phasullia žaunu spraugas;
4 - Bosmina vēžveidīgais ar filtrētu zarnu saturu;
5 - ciliātu Bursaria barības strāva
Vides īpašības lielā mērā nosaka tās iemītnieku pielāgošanās veidus, viņu dzīvesveidu un resursu izmantošanas veidus, veidojot cēloņu-seku attiecību ķēdi. Tādējādi lielais ūdens blīvums padara iespējamu planktona pastāvēšanu, un ūdenī peldošu organismu klātbūtne ir priekšnoteikums filtrācijas veida uztura attīstībai, kurā iespējams arī mazkustīgs dzīvnieku dzīvesveids. Rezultātā veidojas spēcīgs biosfēras nozīmes rezervuāru pašattīrīšanās mehānisms. Tas ietver milzīgu skaitu ūdens organismu, gan bentosa, gan pelaģisko, no vienšūnu vienšūņiem līdz mugurkaulniekiem. Pēc aprēķiniem, viss mērenās joslas ezeros esošais ūdens veģetācijas periodā tiek izlaists caur dzīvnieku filtrēšanas ierīcēm no vairākām līdz desmitiem reižu, un viss Pasaules okeāna tilpums tiek filtrēts vairāku dienu laikā. Filtru padevēju darbības traucējumi dažādu antropogēnas ietekmes dēļ nopietni apdraud ūdeņu tīrības saglabāšanu.
4.2. Dzīves vide zeme-gaiss
Zemes-gaisa vide ir visgrūtākā vides apstākļu ziņā. Dzīvei uz zemes bija nepieciešami tādi pielāgojumi, kas bija iespējami tikai ar pietiekami augstu augu un dzīvnieku organizācijas līmeni.
4.2.1. Gaiss kā vides faktors sauszemes organismiem
Zems gaisa blīvums nosaka tā zemo pacēlumu un nenozīmīgo strīdu. Gaisa vides iemītniekiem ir jābūt savai atbalsta sistēmai, kas atbalsta ķermeni: augiem - dažādiem mehāniskiem audi, dzīvniekiem - cietam vai, daudz retāk, hidrostatiskajam skeletam. Turklāt visi gaisa vides iemītnieki ir cieši saistīti ar zemes virsmu, kas kalpo viņiem pieķeršanās un atbalstam. Apturēta dzīvība gaisā nav iespējama.
Tiesa, daudzi mikroorganismi un dzīvnieki, augu sporas, sēklas, augļi un ziedputekšņi regulāri atrodas gaisā un tos nes gaisa straumes (43. att.), daudzi dzīvnieki spēj aktīvi lidot, taču visās šajās sugās galvenā funkcija to dzīves cikla - vairošanās - tiek veikta uz zemes virsmas. Lielākajai daļai uzturēšanās gaisā asociējas tikai ar iekārtošanos vai medījuma meklēšanu.
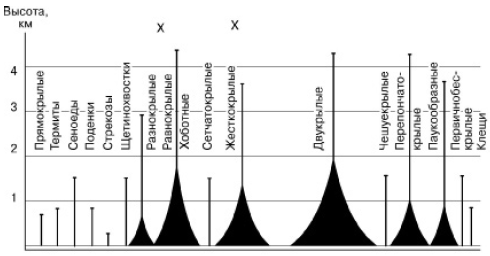
Rīsi. 43. Gaisa planktona posmkāju sadalījums pēc augstuma (saskaņā ar Dajo, 1975)
Zems gaisa blīvums rada zemu pretestību kustībai. Tāpēc daudzi sauszemes dzīvnieki evolūcijas gaitā ir izmantojuši šīs gaisa īpašības ekoloģiskos ieguvumus, iegūstot spēju lidot. 75% visu sauszemes dzīvnieku sugu, galvenokārt kukaiņi un putni, spēj aktīvi lidot, taču skrejlapas ir sastopamas arī starp zīdītājiem un rāpuļiem. Sauszemes dzīvnieki lido galvenokārt ar muskuļu piepūles palīdzību, bet daži var arī slīdēt gaisa straumju dēļ.
Gaisa mobilitātes, atmosfēras apakšējos slāņos esošo gaisa masu vertikālo un horizontālo kustību dēļ iespējama virknes organismu pasīvā lidojuma.
Anemofilija - vecākā augu apputeksnēšanas metode. Visus ģimnosēkļus apputeksnē vējš, un starp segsēkļiem anemofīlie augi veido apmēram 10% no visām sugām.
Anemophilia tiek novērota dižskābarža, bērza, valriekstu, gobu, kaņepju, nātru, kazuarīna, miglas, grīšļa, graudaugu, palmu un daudzu citu ģimenēs. Vēja apputeksnētajiem augiem ir vairāki pielāgojumi, kas uzlabo to putekšņu aerodinamiskās īpašības, kā arī morfoloģiskās un bioloģiskās īpašības, kas nodrošina apputeksnēšanas efektivitāti.
Daudzu augu dzīve ir pilnībā atkarīga no vēja, un ar tā palīdzību tiek veikta pārvietošana. Šāda dubultā atkarība vērojama eglēm, priedēm, papeles, bērziem, gobām, oša kokiem, kokvilnas stiebrzālei, kaķtautēm, saksauliem, juzguniem u.c.
Ir izveidojušās daudzas sugas anemohorija- pārvietošana ar gaisa plūsmu palīdzību. Anemohorija ir raksturīga augu sporām, sēklām un augļiem, vienšūņu cistām, maziem kukaiņiem, zirnekļiem uc Organismus, ko pasīvi nes gaisa straumes, kolektīvi sauc gaisa planktons pēc analoģijas ar planktona iemītniekiem ūdens vidē. Īpaši pielāgojumi pasīvajam lidojumam ir ļoti mazi ķermeņa izmēri, tā laukuma palielināšanās izaugumu dēļ, spēcīga sadalīšana, liela spārnu relatīvā virsma, zirnekļu tīklu izmantošana utt. (44. att.). Arī augu anemohorālajām sēklām un augļiem ir vai nu ļoti mazi izmēri (piemēram, orhideju sēklas), vai arī dažādi pterigoīdiem un izpletņiem līdzīgi piedēkļi, kas palielina to slīdēšanas spēju (45. att.).
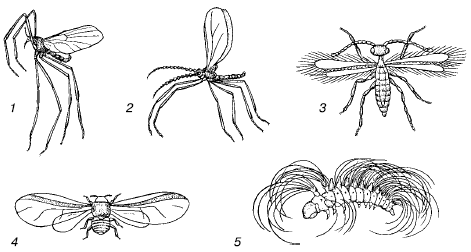
Rīsi. 44. Kukaiņu gaisa transporta pielāgojumi:
1 - ods Cardiocrepis brevirostris;
2 - žultspūslis Porrycordila sp .;
3 - Hymenoptera Anargus fuscus;
4 - Hermes Dreyfusia nordmannianae;
5 - čigānu kodes Lymantria dispar kāpurs
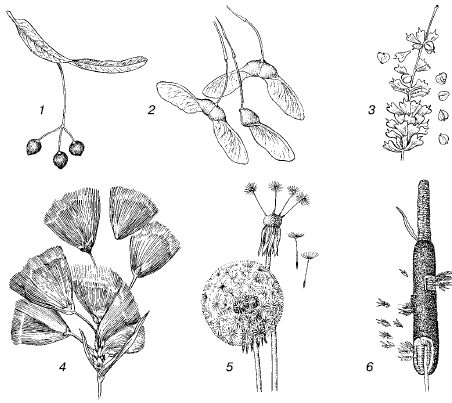
Rīsi. 45. Pielāgojumi vēja transportēšanai augļos un augu sēklās:
1 - liepa Tilia intermedia;
2 - Acer monspessulanum kļava;
3 - bērzs Betula pendula;
4 - kokvilnas zāle Eriophorum;
5 - Pienene Taraxacum officinale;
6 - kaķaste Typha scuttbeworhii
Mikroorganismu, dzīvnieku un augu izplatīšanā galvenā loma ir vertikālām konvekcijas gaisa straumēm un vājam vējam. Spēcīgiem vējiem, vētrām un viesuļvētrām ir arī ievērojama ietekme uz vidi uz sauszemes organismiem.
Zems gaisa blīvums rada salīdzinoši zemu spiedienu uz zemes. Parasti tas ir vienāds ar 760 mm Hg. Art. Palielinoties augstumam, spiediens samazinās. 5800 m augstumā tas ir tikai puse no normas. Zems spiediens var ierobežot sugu izplatību kalnos. Lielākajai daļai mugurkaulnieku dzīves augšējā robeža ir aptuveni 6000 m. Spiediena pazemināšanās izraisa skābekļa piegādes samazināšanos un dzīvnieku dehidratāciju, jo palielinās elpošanas ātrums. Aptuveni tādas pašas ir augstāko augu virzīšanās kalnos robežas. Nedaudz izturīgāki ir posmkāji (atsperes, ērces, zirnekļi), kas sastopami uz ledājiem, virs veģetācijas robežas.
Kopumā visi sauszemes organismi ir daudz stenobātiskāki nekā ūdens organismi, jo parastās spiediena svārstības to vidē veido daļu atmosfēras un pat putniem, kas paceļas lielā augstumā, nepārsniedz 1/3 normālās.
Gaisa gāzes sastāvs. Papildus gaisa fizikālajām īpašībām tā ķīmiskās īpašības ir ārkārtīgi svarīgas sauszemes organismu pastāvēšanai. Gaisa gāzu sastāvs atmosfēras virsmas slānī ir diezgan viendabīgs attiecībā pret galveno komponentu saturu (slāpeklis - 78,1%, skābeklis - 21,0, argons - 0,9, oglekļa dioksīds - 0,035 tilpuma%), jo augsta gāzu difūzijas jauda un pastāvīga konvekcijas un vēja plūsmu sajaukšanās. Tomēr dažādiem gāzveida, pilienu-šķidruma un cieto (putekļu) daļiņu piemaisījumiem, kas no vietējiem avotiem nonāk atmosfērā, var būt nozīmīga vides nozīme.
Augstais skābekļa saturs veicināja sauszemes organismu metabolisma palielināšanos salīdzinājumā ar primārajiem ūdens organismiem. Tieši sauszemes vidē, pamatojoties uz augstu oksidatīvo procesu efektivitāti organismā, radās dzīvnieku homeotermija. Skābeklis, pateicoties tā pastāvīgi augstajam saturam gaisā, nav faktors, kas ierobežo dzīvību sauszemes vidē. Tikai vietām, konkrētos apstākļos, īslaicīgi ir deficīts, piemēram, trūdošu augu atlieku uzkrājumos, graudu, miltu krājumos u.c.
Oglekļa dioksīda saturs noteiktās virszemes gaisa slāņa vietās var atšķirties diezgan ievērojamās robežās. Piemēram, ja lielu pilsētu centrā nav vēja, tā koncentrācija palielinās desmitkārtīgi. Oglekļa dioksīda satura diennakts izmaiņas virsmas slāņos, kas saistītas ar augu fotosintēzes ritmu, ir dabiskas. Sezonālas izraisa dzīvo organismu, galvenokārt augsnes mikroskopiskās populācijas, elpošanas intensitātes izmaiņas. Paaugstināts gaisa piesātinājums ar oglekļa dioksīdu notiek vulkāniskās aktivitātes zonās, termālo avotu un citu šīs gāzes pazemes izplūdes vietu tuvumā. Oglekļa dioksīds ir toksisks augstā koncentrācijā. Dabā šādas koncentrācijas ir reti sastopamas.
Dabā galvenais oglekļa dioksīda avots ir tā sauktā augsnes elpošana. Augsnes mikroorganismi un dzīvnieki elpo ļoti intensīvi. Oglekļa dioksīds izkliedējas no augsnes atmosfērā, īpaši lietus laikā. Daudz no tā izdala vidēji mitras, labi uzsildītas un ar organiskām atliekām bagātas augsnes. Piemēram, dižskābarža meža augsne izdala CO 2 no 15 līdz 22 kg / ha stundā, bet neapaugļota smilšaina augsne - tikai 2 kg / ha.
Mūsdienu apstākļos cilvēka darbība, lai sadedzinātu fosilā kurināmā rezerves, ir kļuvusi par spēcīgu papildu CO 2 daudzuma avotu atmosfērā.
Gaisa slāpeklis lielākajai daļai sauszemes vides iemītnieku ir inerta gāze, taču virkne prokariotu organismu (mezglu baktērijas, azotobaktērijas, klostridijas, zilaļģes u.c.) spēj to saistīt un iesaistīt bioloģiskajā. apgrozībā.

Rīsi. 46. Kalna nogāze ar iznīcinātu veģetāciju apkārtējo rūpniecības uzņēmumu sēra dioksīda emisiju dēļ
Vietējie piemaisījumi, kas nonāk gaisā, var būtiski ietekmēt arī dzīvos organismus. Īpaši tas attiecas uz indīgām gāzveida vielām - metānu, sēra oksīdu, oglekļa monoksīdu, slāpekļa oksīdu, sērūdeņradi, hlora savienojumiem, kā arī putekļu, kvēpu u.c. daļiņām, kas aizsprosto gaisu industriālajos rajonos. Mūsdienu galvenais atmosfēras ķīmiskā un fizikālā piesārņojuma avots ir antropogēns: dažādu rūpniecības uzņēmumu un transporta darbs, augsnes erozija uc Piemēram, sēra oksīds (SO 2) ir indīgs augiem pat koncentrācijā no vienas piecdesmit tūkstošdaļas. līdz vienai miljonajai daļai no gaisa tilpuma. Ap industriālajiem centriem, kas piesārņo atmosfēru ar šo gāzi, iet bojā gandrīz visa veģetācija (46. att.). Dažas augu sugas ir īpaši jutīgas pret SO 2 un kalpo kā jutīgs indikators tā uzkrāšanai gaisā. Piemēram, daudzi ķērpji iet bojā pat ar sēra oksīda pēdām apkārtējā atmosfērā. To klātbūtne mežos ap lielajām pilsētām liecina par augstu gaisa tīrību. Izvēloties sugas ainavu veidošanai apdzīvotās vietās, tiek ņemta vērā augu izturība pret gaisa piemaisījumiem. Dūmu jutīgas, piemēram, egle un priede, kļava, liepa, bērzs. Visizturīgākās ir tūjas, Kanādas papele, Amerikas kļava, plūškoks un daži citi.
4.2.2. Augsne un reljefs. Zemes-gaisa vides laikapstākļi un klimatiskie apstākļi
Edafiskie vides faktori. Augsnes un reljefa īpašības ietekmē arī sauszemes organismu, galvenokārt augu, dzīves apstākļus. Zemes virsmas īpašības, kurām ir ekoloģiska ietekme uz tās iedzīvotājiem, vieno nosaukums edafiskie vides faktori (no grieķu "edaphos" - pamatne, augsne).
Augu sakņu sistēmas raksturs ir atkarīgs no hidrotermālā režīma, aerācijas, augsnes sastāva, sastāva un struktūras. Piemēram, koku sugu (bērzs, lapegle) sakņu sistēmas teritorijās ar mūžīgo sasalumu atrodas seklā dziļumā un izplatās platumā. Tur, kur nav mūžīgā sasaluma, to pašu augu sakņu sistēmas ir mazāk izplatītas un iekļūst dziļāk. Daudzos stepju augos saknes var iegūt ūdeni no liela dziļuma, tajā pašā laikā tām ir arī daudzas virszemes saknes augsnes trūdvielu horizontā, no kurienes augi uzņem minerālbarības elementus. Uz ūdens piesātinātas, slikti gāzētas augsnes mangrovju biezokņos daudzām sugām ir īpašas elpošanas saknes - pneimatofori.
Saistībā ar dažādām augsnes īpašībām var izdalīt vairākas ekoloģiskās augu grupas.
Tātad, atkarībā no reakcijas uz augsnes skābumu, tos izšķir: 1) acidofils sugas - aug skābās augsnēs, kuru pH ir mazāks par 6,7 (sfagnu purva augi, baltspārni); 2) neitrofīls - gravitēt uz augsnēm ar pH 6,7–7,0 (lielākā daļa kultivēto augu); 3) bazifilisks- augt pie pH vairāk nekā 7,0 (mordovnik, meža anemone); 4) vienaldzīgs - var augt augsnēs ar dažādām pH vērtībām (maijlilija, aitu auzene).
Attiecībā uz augsnes bruto sastāvu ir: 1) oligotrofisks augi, kas ir apmierināti ar nelielu daudzumu oša elementu (parastā priede); 2) eitrofisks, tiem, kam nepieciešams liels skaits oša elementu (ozols, sārtais, daudzgadīgais meža audzētājs); 3) mezotrofisks, kam nepieciešams mērens oša elementu daudzums (parastā egle).
Nitrofīli- augi, kas dod priekšroku ar slāpekli bagātām augsnēm (nātre).
Sāls augsnes augi veido grupu halofīti(saltros, sarsazāns, kokpeks).
Dažas augu sugas ir ierobežotas ar dažādiem substrātiem: petrofīti augt akmeņainās augsnēs, un psammofīti apdzīvot irdenas smiltis.
Reljefs un zemes raksturs ietekmē dzīvnieku īpašo kustību. Piemēram, pārnadžiem, strausiem un dumpēm, kas dzīvo atklātās vietās, ir nepieciešama stabila augsne, lai palielinātu atbaidīšanu, ātri skrienot. Uz irdenām smiltīm dzīvojošām ķirzakām pirkstus norobežo ragveida zvīņu bārkstis, kas palielina balsta virsmu (47. att.). Sauszemes iedzīvotājiem, kas rok bedrītes, blīvas augsnes ir nelabvēlīgas. Augsnes raksturs dažos gadījumos ietekmē sauszemes dzīvnieku izplatību, ierakšanu, ierakšanu zemē, lai izvairītos no karstuma vai plēsējiem, vai olu ievietošanu augsnē utt.
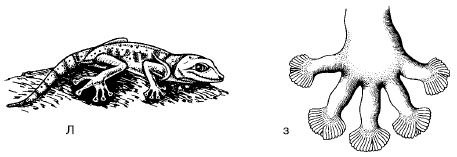
Rīsi. 47. Vēdereņgekons - Sahāras smilšu iemītnieks: A - vēdekļpēdu gekons; B - gekona kāja
Laikapstākļu pazīmes. Dzīves apstākļi zemes-gaisa vidē ir sarežģīti, turklāt laikapstākļu izmaiņas. Laikapstākļi - tas ir nepārtraukti mainīgs atmosfēras stāvoklis zemes virsmas tuvumā līdz aptuveni 20 km augstumam (troposfēras robeža). Laikapstākļu mainīgums izpaužas tādu vides faktoru kā gaisa temperatūra un mitrums, mākoņainība, nokrišņi, vēja stiprums un virziens uc sauszemes organismu kombinācijas pastāvīgā variācijā. Laikapstākļi ūdens iemītnieku dzīvi ietekmē daudz mazākā mērā un tikai virszemes slāņu populāciju.
Apgabala klimats. Raksturo ilgstošu laikapstākļu režīmu apgabala klimats. Klimata jēdziens ietver ne tikai meteoroloģisko parādību vidējās vērtības, bet arī to ikgadējās un ikdienas izmaiņas, novirzes no tā un to biežumu. Klimatu nosaka apgabala ģeogrāfiskie apstākļi.
Klimata zonālo daudzveidību sarežģī musonu vēju ietekme, ciklonu un anticiklonu izplatība, kalnu grēdu ietekme uz gaisa masu kustību, attāluma pakāpe no okeāna (kontinentalitāte), kā arī daudzi citi lokāli faktori. Kalnos ir klimatiskais zonējums, kas daudzos aspektos ir līdzīgs zonu maiņai no zemiem uz augstiem platuma grādiem. Tas viss rada ārkārtīgi daudzveidīgus dzīves apstākļus uz sauszemes.
Lielākajai daļai sauszemes organismu, īpaši mazajiem, svarīgs ir ne tik daudz reģiona klimats, cik apstākļi to tiešā dzīvotnē. Ļoti bieži vietējie vides elementi (reljefs, ekspozīcija, veģetācija u.c.) maina temperatūras, mitruma, gaismas, gaisa kustības režīmu noteiktā teritorijā tā, ka tas būtiski atšķiras no apgabala klimatiskajiem apstākļiem. Tādas lokālās klimata modifikācijas, kas attīstās virszemes gaisa slānī, sauc mikroklimats. Mikroklimats katrā zonā ir ļoti dažāds. Var izdalīt patvaļīgi mazu platību mikroklimatus. Piemēram, ziedu vainagā tiek izveidots īpašs režīms, ko izmanto tur mītošie kukaiņi. Plaši zināmas temperatūras, gaisa mitruma un vēja spēka atšķirības atklātā kosmosā un mežā, zālē un virs kailas augsnes, ziemeļu un dienvidu atsegumu nogāzēs uc Īpašs stabils mikroklimats veidojas urvos, ligzdās. , ieplakas, alas un citas.slēgtas vietas.
Nokrišņi. Papildus ūdens apgādei un mitruma uzglabāšanai tiem var būt vēl viena ekoloģiska nozīme. Tādējādi spēcīgas lietusgāzes vai krusa dažkārt mehāniski ietekmē augus vai dzīvniekus.
Sniega segas ekoloģiskā loma ir īpaši daudzveidīga. Diennakts temperatūras svārstības sniega biezumā iekļūst tikai līdz 25 cm, dziļāk temperatūra gandrīz nemainās. Ar -20-30 ° C salnām zem 30-40 cm sniega kārtas temperatūra ir tikai nedaudz zem nulles. Dziļa sniega sega pasargā atjaunošanās pumpurus, aizsargā augu zaļās daļas no sasalšanas; daudzas sugas nokļūst zem sniega, nenometot lapotni, piemēram, apmatojuma miza, Veronica officinalis, šķeltne u.c.
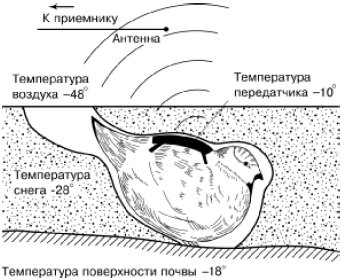
Rīsi. 48. Lazdu rubeņu temperatūras režīma telemetriskā pētījuma shēma, kas atrodas sniegotā bedrē (pēc A. V. Andrejeva, A. V. Krečmara, 1976)
Mazie sauszemes dzīvnieki ziemā piekopj aktīvu dzīvesveidu, zem sniega un tā biezumā izklājot veselas eju galerijas. Vairākām sugām, kas barojas ar sniegotu veģetāciju, raksturīga vienmērīga vairošanās ziemā, kas novērota, piemēram, lemmingiem, meža un dzeltenkakla pelēm, virknei pelēm, ūdensžurkām u.c. 48. att.).
Ziemas sniega sega neļauj lieliem dzīvniekiem iegūt barību. Daudzi nagaiņi (ziemeļbrieži, mežacūkas, muskusa vērši) ziemā barojas tikai ar sniegotu veģetāciju, un dziļa sniega sega un īpaši cieta garoza uz tās virsmas, kas parādās ledū, nolemj tos barības trūkumam. Nomadu liellopu audzēšanas laikā pirmsrevolūcijas Krievijā notika milzīga katastrofa dienvidu reģionos. džuta - masveida mājlopu nāve ledus rezultātā, atņemot dzīvniekiem barību. Pārvietošanās pa irdenu dziļu sniegu arī dzīvniekiem ir apgrūtināta. Lapsas, piemēram, sniegotās ziemās dod priekšroku vietām mežā zem blīvām eglēm, kur sniega kārta ir plānāka, un gandrīz nekad neiet uz atklātām pļavām un mežmalām. Sniega segas dziļums var ierobežot sugu ģeogrāfisko izplatību. Piemēram, īstie brieži neiekļūst ziemeļos uz tiem apgabaliem, kur sniega biezums ziemā ir lielāks par 40-50 cm.
Sniega segas baltums atklāj tumšos dzīvniekus. Sezonālas krāsas maiņas rašanās baltajām un tundras irbēm, baltajam zaķim, erminam, zebiekstei, polārlapsai, acīmredzami liela nozīme bija izvēlei, lai maskētu fona krāsu. Komandieru salās kopā ar baltajiem ir daudz zilo lapsu. Pēc zoologu novērojumiem pēdējie uzturas galvenokārt pie tumšiem akmeņiem un neaizsalstoša sērfošanas josla, bet baltie dod priekšroku vietām ar sniega segu.
4.3. Augsne kā biotops
4.3.1. Augsnes īpašības
Augsne ir irdens plāns zemes virsmas slānis, kas saskaras ar gaisu. Neskatoties uz nenozīmīgo biezumu, šim Zemes apvalkam ir izšķiroša nozīme dzīvības izplatībā. Augsne nav tikai cieta viela, tāpat kā lielākā daļa litosfēras iežu, bet gan sarežģīta trīsfāžu sistēma, kurā cietās daļiņas ieskauj gaiss un ūdens. Tas ir caurstrāvots ar dobumiem, kas piepildīti ar gāzu un ūdens šķīdumu maisījumu, un tāpēc tajā veidojas ārkārtīgi daudzveidīgi apstākļi, kas ir labvēlīgi daudzu mikro- un makroorganismu dzīvībai (49. att.). Augsnē temperatūras svārstības tiek izlīdzinātas salīdzinājumā ar virszemes gaisa slāni, un gruntsūdeņu klātbūtne un nokrišņu iekļūšana veido mitruma rezerves un nodrošina mitruma režīmu starpposmā starp ūdens un sauszemes vidi. Augsnē koncentrējas organisko un minerālvielu krājumi, ko piegādā mirstoša veģetācija un dzīvnieku līķi. Tas viss nosaka augsto piesātinājumu ar dzīvību.
Sauszemes augu sakņu sistēmas ir koncentrētas augsnē (50. att.).
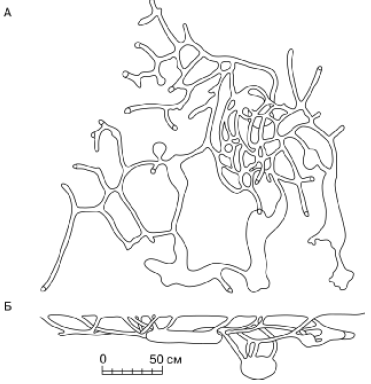
Rīsi. 49. Brandta straumes pazemes ejas: A - skats no augšas; B - sānskats
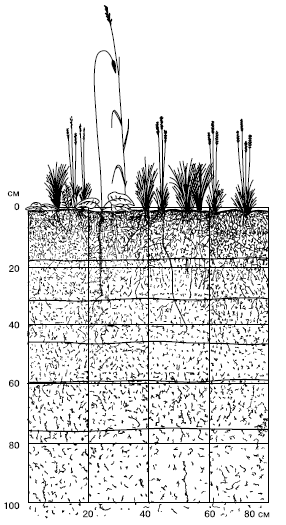
Rīsi. 50. Sakņu izvietošana stepju chernozem augsnē (pēc M.S.Shalyt, 1950)
Vidēji 1 m 2 augsnes slāņa veido vairāk nekā 100 miljardus vienšūņu šūnu, miljoniem rotiferu un tardigradu, desmitiem miljonu nematožu, desmitiem un simtiem tūkstošu ērču un kolembolānu, tūkstošiem citu posmkāju, desmitiem tūkstošu enhitreīdas, desmitiem un simtiem slieku, mīkstmiešu un citu bezmugurkaulnieku ... Turklāt 1 cm 2 augsnes satur desmitiem un simtiem miljonu baktēriju, mikroskopisku sēņu, aktinomicītu un citu mikroorganismu. Apgaismotajos virsmas slāņos katrā gramā dzīvo simtiem tūkstošu zaļo, dzeltenzaļo, kramaļģu un zilaļģu fotosintēzes šūnu. Dzīvie organismi ir tikpat raksturīgi augsnei kā tās nedzīvās sastāvdaļas. Tāpēc V.I.Vernadskis augsni klasificēja kā vienu no bioinertajiem dabas ķermeņiem, uzsverot tās dzīves bagātību un nesaraujamo saistību ar to.
Apstākļu neviendabīgums augsnē visspilgtāk izpaužas vertikālā virzienā. Vairāki svarīgākie ekoloģiskie faktori, kas ietekmē augsnes iemītnieku dzīvi, krasi mainās līdz ar dziļumu. Pirmkārt, tas attiecas uz augsnes struktūru. Tajā tiek izdalīti trīs galvenie horizonti, kas atšķiras pēc morfoloģiskajām un ķīmiskajām īpašībām: 1) augšējais trūdvielu akumulatīvais horizonts A, kurā uzkrājas un transformējas organiskās vielas un no kura daļu savienojumu novada mazgāšanas ūdeņi; 2) ieskalošanās horizonts jeb iluviālais B, kur nosēžas un transformējas no augšas izskalotās vielas, un 3) pamatiežu jeb C horizontu, kura materiāls pārvēršas augsnē.
Katrā horizontā izšķir vairāk frakcionētu slāņu, kas arī ļoti atšķiras pēc īpašībām. Piemēram, mērenā joslā zem skujkoku vai jauktiem mežiem, horizonts A sastāv no pakaišiem (A 0)- irdenas augu atlieku uzkrāšanās slānis, tumšas krāsas humusa slānis (A 1), kurā organiskas izcelsmes daļiņas sajauktas ar minerālu un podzolisko slāni (A 2)- pelnu pelēkā krāsā, kurā dominē silīcija savienojumi, un visas šķīstošās vielas tiek ieskalotas augsnes profila dziļumā. Gan šo slāņu struktūra, gan ķīmija ir ļoti atšķirīga, un tāpēc augu saknes un augsnes iemītnieki, pārvietojoties tikai dažus centimetrus uz augšu vai uz leju, nonāk dažādos apstākļos.
Dobumu izmēri starp augsnes daļiņām, kas piemērotas dzīvnieku apdzīvošanai, parasti strauji samazinās līdz ar dziļumu. Piemēram, pļavu augsnēs dobumu vidējais diametrs 0–1 cm dziļumā ir 3 mm, 1–2 cm - 2 mm, bet 2–3 cm dziļumā - tikai 1 mm; dziļāk augsnes poras ir vēl mazākas. Augsnes blīvums mainās arī līdz ar dziļumu. Irdenākie ir organisko vielu saturošie slāņi. Šo slāņu porainību nosaka tas, ka organiskās vielas salipina kopā minerālu daļiņas lielākos agregātos, starp kuriem palielinās dobumu tilpums. Visblīvākais parasti ir iluviālais horizonts V, sacementē tajā izskalotās koloidālās daļiņas.
Mitrums augsnē ir dažādos stāvokļos: 1) saistīts (higroskopisks un plēves) stingri notur augsnes daļiņu virsma; 2) kapilārs aizņem nelielas poras un var pārvietoties pa tām dažādos virzienos; 3) gravitācija aizpilda lielākus tukšumus un gravitācijas ietekmē lēnām sūcas lejup; 4) tvaiki atrodas augsnes gaisā.
Ūdens saturs dažādās augsnēs un dažādos laikos nav vienāds. Ja ir pārāk daudz gravitācijas mitruma, tad augsnes režīms ir tuvs ūdenstilpju režīmam. Sausā augsnē paliek tikai saistīts ūdens, un apstākļi tuvojas sauszemes apstākļiem. Tomēr pat sausākajās augsnēs gaiss ir mitrāks nekā virs zemes, tāpēc augsnes iemītnieki ir daudz mazāk pakļauti izžūšanas draudiem nekā virspusē.
Augsnes gaisa sastāvs ir mainīgs. Ar dziļumu tajā strauji samazinās skābekļa saturs un palielinās oglekļa dioksīda koncentrācija. Sadalīšanās organisko vielu klātbūtnes dēļ augsnē augsnes gaiss var saturēt augstu toksisku gāzu koncentrāciju, piemēram, amonjaku, sērūdeņradi, metānu u.c. Kad augsne ir appludināta vai notiek intensīva augu atlieku puve, var rasties pilnīgi anaerobi apstākļi. rodas vietām.
Pļaušanas temperatūras svārstības tikai pie augsnes virsmas. Šeit tie var būt pat stiprāki nekā virszemes gaisa slānī. Taču ar katru centimetru dziļāk diennakts un sezonālās temperatūras izmaiņas kļūst arvien mazākas un 1–1,5 m dziļumā praktiski vairs nav izsekojamas (51. att.).
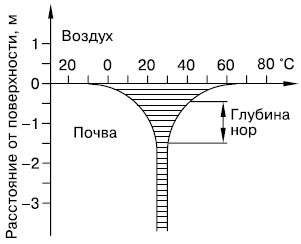
Rīsi. 51. Augsnes temperatūras ikgadējo svārstību samazināšanās līdz ar dziļumu (saskaņā ar K. Schmidt-Nilsson, 1972). Aizēnotā daļa ir gada temperatūras svārstību diapazons
Visas šīs īpašības noved pie tā, ka, neraugoties uz lielo vides apstākļu neviendabīgumu augsnē, tā darbojas kā diezgan stabila vide, īpaši mobilajiem organismiem. Stāvais temperatūras un mitruma gradients augsnes profilā ļauj augsnes dzīvniekiem nodrošināt sev piemērotu ekoloģisko situāciju, veicot nelielas kustības.
4.3.2. Augsnes iedzīvotāji
Augsnes neviendabīgums noved pie tā, ka dažāda lieluma organismiem tā darbojas kā atšķirīga vide. Mikroorganismiem milzīga augsnes daļiņu kopējā virsma ir īpaši svarīga, jo lielākā daļa mikrobu populācijas ir uz tām adsorbēta. Augsnes vides sarežģītība rada visdažādākos apstākļus dažādām funkcionālajām grupām: aerobiem un anaerobiem, organisko un minerālu savienojumu patērētājiem. Mikroorganismu izplatību augsnē raksturo neliels fokuss, jo pat vairāku milimetru garumā var mainīties dažādas ekoloģiskās zonas.
Nelieliem augsnes dzīvniekiem (52., 53. att.), kas apvienoti zem nosaukuma mikrofauna (vienšūņi, rotifers, tardigrades, nematodes uc), augsne ir mikrorezervuāru sistēma. Būtībā tie ir ūdens organismi. Tie dzīvo augsnes porās, kas piepildītas ar gravitācijas vai kapilāru ūdeni, un daļa no to dzīves, tāpat kā mikroorganismi, var adsorbēties uz daļiņu virsmas plānos plēves mitruma slāņos. Daudzas no šīm sugām dzīvo arī parastās ūdenstilpēs. Tomēr augsnes formas ir daudz mazākas nekā saldūdens formas, un turklāt tās izceļas ar spēju ilgstoši palikt ensistētā stāvoklī, gaidot nelabvēlīgus periodus. Kamēr saldūdens amēbas ir 50-100 mikronu lielas, augsnes - tikai 10-15. Flagelātu pārstāvji ir īpaši mazi, bieži vien tikai 2–5 µm. Augsnes skropstiņi ir arī pundura izmēra un turklāt var ievērojami mainīt ķermeņa formu.
Rīsi. 52. Amēba, kas barojas ar baktērijām uz trūdošām meža zemsedzes lapām

Rīsi. 53. Augsnes mikrofauna (pēc V. Dangera, 1974):
1–4 - flagellate; 5–8 - kaila amēba; 9-10 - gliemežvāku amēba; 11–13 - ciliāti; 14–16 - apaļie tārpi; 17–18 - rotiferi; 19–20 – tardigrades
Nedaudz lielākiem dzīvniekiem, kas elpo gaisu, augsne šķiet kā mazu alu sistēma. Šādi dzīvnieki ir apvienoti zem nosaukuma mezofauna (54. att.). Augsnes mezofaunas pārstāvju izmēri ir no desmitdaļām līdz 2–3 mm. Šajā grupā galvenokārt ietilpst posmkāji: daudzas ērču grupas, primārie kukaiņi bez spārniem (kolemboli, protora, divastes), nelielas spārnotu kukaiņu sugas, simtkāji simfilas uc Viņiem nav īpašu instrumentu rakšanai. Viņi rāpo gar augsnes dobumu sienām ar savu ekstremitāšu vai tārpiem līdzīgu tārpu palīdzību. Ar ūdens tvaikiem piesātinātais augsnes gaiss ļauj elpot caur apvalku. Daudzām sugām nav trahejas sistēmas. Šādi dzīvnieki ir ļoti jutīgi pret izžūšanu. Viņiem galvenais glābiņš no gaisa mitruma svārstībām ir pārcelšanās uz iekšzemi. Bet migrācijas iespēju caur augsnes dobumiem dziļumā ierobežo straujš poru diametra samazinājums, tāpēc pārvietošanās pa augsnes akām ir pieejama tikai mazākajām sugām. Lielākiem mezofaunas pārstāvjiem ir daži pielāgojumi, kas ļauj izturēt īslaicīgu augsnes gaisa mitruma samazināšanos: aizsargzvīņas uz ķermeņa, daļēja apvalku necaurlaidība, ciets biezsienu apvalks ar epikutikulu kombinācijā ar primitīvu trahejas sistēmu, kas nodrošina elpošana.
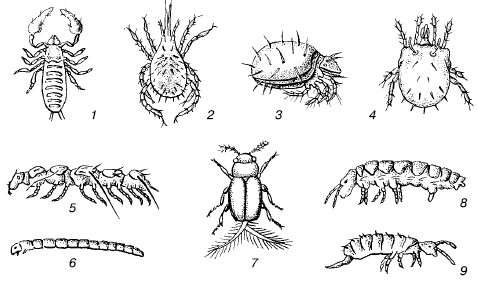
Rīsi. 54. Augsņu mezofauna (bez W. Danger, 1974):
1 - viltus skorions; 2 - din jaunu lāpu; 3–4 bruņu ērces; 5 - pauroiod simtkājis; 6 - Chironomid moskītu kāpurs; 7 - vabole no šī. Ptiliidae; 8–9 kolembolāni
Mezofaunas pārstāvji pārdzīvo augsnes applūdināšanas periodus ar ūdeni gaisa burbuļos. Gaiss tiek aizturēts ap dzīvnieku ķermeni, pateicoties to nesamitrinošajiem apvalkiem, kas tiek apgādāti arī ar matiem, zvīņām utt. Gaisa burbulis kalpo kā sava veida "fiziskā žauna" mazam dzīvniekam. Elpošana notiek, jo skābeklis no apkārtējā ūdens izkliedējas gaisa slānī.
Mikro- un mezofaunas pārstāvji spēj izturēt ziemas augsnes sasalšanu, jo lielākā daļa sugu nevar nolaisties no slāņiem, kas pakļauti negatīvai temperatūrai.
Lielākus augsnes dzīvniekus, kuru ķermeņa izmērs ir no 2 līdz 20 mm, sauc par pārstāvjiem makrofauna (55. att.). Tie ir kukaiņu kāpuri, tūkstoškājai, enhitreīdas, sliekas uc Viņiem augsne ir blīva vide, kas nodrošina ievērojamu mehānisko pretestību kustoties. Šīs salīdzinoši lielās formas augsnē pārvietojas, vai nu paplašinot dabiskās akas, atstumjot augsnes daļiņas, vai arī izrokot jaunas ejas. Abi kustības veidi atstāj iespaidu uz dzīvnieku ārējo struktūru.
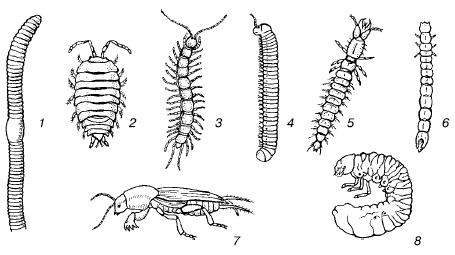
Rīsi. 55. Augsņu makrofauna (bez W. Danger, 1974):
1 - slieka; 2 – koka utis; 3 – simtkāji; 4 – divkājains simtkājis; 5 - zemes vaboļu kāpurs; 6 – riekstkoka kāpurs; 7 – lācis; 8 - grumbu kāpurs
Spēja pārvietoties pa plānām akām, gandrīz neizmantojot rakšanu, ir raksturīga tikai sugām, kuru ķermenis ir ar mazu šķērsgriezumu, kas spēj stipri saliekties līkumotās ejās (simtkāji - kauleņi un ģeofīli). Atgrūžot augsnes daļiņas ķermeņa sieniņu spiediena ietekmē, pārvietojas sliekas, garkājainu odu kāpuri u.c.. Nofiksējuši aizmugurējo galu, tie retāk un pagarina priekšējo galu, iekļūstot šaurās augsnes plaisās, pēc tam nofiksē. ķermeņa priekšējo daļu un palielināt tās diametru. Tajā pašā laikā palielinātajā zonā, pateicoties muskuļu darbam, tiek izveidots spēcīgs nesaspiežamā intrakavitārā šķidruma hidrauliskais spiediens: tārpiem - celomisko maisiņu saturs, bet tipulīdos - hemolimfa. Spiediens caur ķermeņa sienām tiek pārnests uz augsni, un tādējādi dzīvnieks paplašina aku. Tajā pašā laikā aiz muguras paliek atvērta eja, kas draud ar pastiprinātu iztvaikošanu un plēsēju vajāšanu. Daudzām sugām ir izstrādāti pielāgojumi ekoloģiski izdevīgākam pārvietošanās veidam augsnē - rakšanai ar ejas aizsērēšanu. Rakšana tiek veikta, irdinot un grābjot augsnes daļiņas. Šim nolūkam dažādu kukaiņu kāpuri izmanto galvas priekšējo galu, apakšžokļus un priekškājas, kas paplašinātas un pastiprinātas ar biezu hitīna slāni, muguriņas un izaugumiem. Ķermeņa aizmugurējā galā attīstās ierīces spēcīgai fiksācijai - izvelkami balsti, zobi, āķi. Lai aizvērtu eju pēdējos segmentos, vairākām sugām ir īpaša nospiesta platforma, ko ierāmē hitīna malas vai zobi, sava veida ķerra. Līdzīgas platformas veidojas elytras aizmugurē un mizgraužos, kuras arī izmanto, lai aizsprostotu ejas ar urbja miltiem. Aizverot eju aiz viņiem, dzīvnieki - augsnes iemītnieki pastāvīgi atrodas slēgtā kamerā, kas piesātināta ar viņu pašu ķermeņa tvaikiem.
Lielākajai daļai šīs ekoloģiskās grupas sugu gāzu apmaiņa tiek veikta ar specializētu elpošanas orgānu palīdzību, bet tajā pašā laikā to papildina gāzu apmaiņa caur integumentiem. Ir iespējama pat tikai ādas elpošana, piemēram, sliekām, enchitreidiem.
Ieraktie dzīvnieki var atstāt slāņus, kur veidojas nelabvēlīga vide. Sausumā un ziemā tie koncentrējas dziļākos slāņos, parasti dažus desmitus centimetru no virsmas.
Megafauna augsnes ir lieli racēji, galvenokārt zīdītāji. Daudzas sugas visu savu dzīvi pavada augsnē (kurmju žurkas, kurmju kurmji, zokori, Eirāzijas kurmji, zelta kurmji
Āfrika, Austrālijas marsupial kurmji utt.). Viņi augsnē ieliek veselas tuneļu un caurumu sistēmas. Šo dzīvnieku ārējais izskats un anatomiskās īpatnības atspoguļo to pielāgošanās spēju pazemes dzīvesveidam. Viņiem ir mazattīstītas acis, kompakts, slīdošs ķermenis ar īsu kaklu, īsa bieza kažokāda, spēcīgas zarnās ar spēcīgiem nagiem. Kurmji un kurmju žurkas atbrīvo zemi ar priekšzobiem. Pie augsnes megafaunas jāpieskaita arī lielie oligošeti, īpaši tropos un dienvidu puslodē mītošie Megascolecidae dzimtas pārstāvji. Lielākais no tiem, Austrālijas Megascolides australis, sasniedz 2,5 un pat 3 m garumu.
Papildus pastāvīgajiem augsnes iemītniekiem starp lielajiem dzīvniekiem var izdalīt lielu ekoloģisko grupu alu iemītnieki (goferi, murkšķi, jerboas, truši, āpši utt.). Tie barojas uz virsmas, bet vairojas, pārziemo, atpūšas, izbēg no briesmām augsnē. Vairāki citi dzīvnieki izmanto savas alas, atrodot tajās labvēlīgu mikroklimatu un patvērumu no ienaidniekiem. Nornikiem ir strukturālas iezīmes, kas raksturīgas sauszemes dzīvniekiem, taču tiem ir vairāki pielāgojumi, kas saistīti ar ierakumu dzīvesveidu. Piemēram, āpšiem ir gari nagi un spēcīgi priekškāju muskuļi, šaura galva un mazas ausis. Trušiem, salīdzinot ar zaķiem, kuri nerok caurumus, ausis un pakaļkājas ir manāmi saīsinātas, galvaskauss stiprāks, attīstītāki apakšdelmu kauli un muskuļi utt.
Vairākām ekoloģiskām īpašībām augsne ir starpvide starp ūdeni un zemi. Augsni ūdens videi tuvina tās temperatūras režīms, zemais skābekļa saturs augsnes gaisā, tās piesātinājums ar ūdens tvaikiem un ūdens klātbūtne citās formās, sāļu un organisko vielu klātbūtne augsnes šķīdumos, spēja pārvietoties trīs dimensijās.
Augsni gaisam tuvina augsnes gaisa klātbūtne, izžūšanas draudi augšējos horizontos un diezgan krasas virsmas slāņu temperatūras režīma izmaiņas.
Augsnes kā dzīvnieku dzīvotnes vidējās ekoloģiskās īpašības liecina, ka augsnei bija īpaša loma dzīvnieku pasaules evolūcijā. Daudzām grupām, jo īpaši posmkājiem, augsne kalpoja kā vide, caur kuru sākotnēji ūdens iemītnieki varēja pāriet uz sauszemes dzīvesveidu un iekarot zemi. Šo posmkāju evolūcijas ceļu pierādīja M. S. Giļarova (1912–1985) darbi.
4.4. Dzīvie organismi kā biotops
Daudzu veidu heterotrofie organismi visu mūžu vai dzīves cikla daļu dzīvo citās dzīvās būtnēs, kuru ķermeņi tiem kalpo kā vide, kas pēc īpašībām būtiski atšķiras no ārējās.
Rīsi. 56. Laputu jātnieks
Rīsi. 57. Izgriezt žulti uz dižskābarža lapas ar žaunu kāpuru Mikiola fagi