Митохондрийн дотоод бүтэц гэж нэрлэдэг. Митохондри ба пластидын бүтэц, үүрэг
Митохондриа
Амьтны эд эсийн эсэд митохондриа 1882 онд, харин ургамалд зөвхөн 1904 онд (усны сараана цэцгийн антритууд) олдсон байна. Фракцыг центрифугалах замаар фракцыг салгаж, цэвэрлэсний дараа биологийн функцийг байгуулах боломжтой байв. Тэд 70% уураг, 30% орчим липид, бага хэмжээний РНХ, ДНХ, А, В 6, В 12, К, Е витамин, фолийн болон пантотен хүчил, рибофлавин, янз бүрийн фермент агуулдаг. Митохондри нь давхар мембрантай, гаднах хэсэг нь органеллийг цитоплазмаас тусгаарладаг ба дотор нь crista-ийн гадагш ургадаг. Мембраны хоорондох зайг бүхэлд нь матрицаар дүүргэдэг (Зураг 13).
Митохондрийн үндсэн үүрэг бол эсийн амьсгалын үйл ажиллагаанд оролцдог. Амьсгалын үед митохондрийн үүрэг 1950-1951 онд байгуулагдсан. Кребсийн мөчлөгийн цогц ферментийн систем нь гаднах мембран дээр төвлөрдөг. Амьсгалын субстратыг исэлдүүлэх үед энерги ялгарч, үүссэн ADP молекулуудад, голчлон ATP, кристацид үүсдэг исэлдэлтийн фосфоржилтын үед үүсдэг. Макроэлементийн нэгдэлд хадгалагдсан энергийг ирээдүйд эсийн бүх хэрэгцээг хангахад ашигладаг.
Эсийн доторх митохондри үүсэх нь бичил биетүүдээс тасралтгүй явагддаг бөгөөд тэдгээрийн илрэл нь эсийн мембраны бүтцийг ялгахтай холбоотой байдаг. Тэдгээрийг хувааж, нахиалах замаар тэдгээрийг эс дотор сэргээж болно. Митохондриа нь удаан эдэлгээтэй биш, дундаж наслалт 5-10 хоног байдаг.
Митохондриа бол "хүч" эсийн станц юм. Тэд энерги нь "аккумулятор" - ATP молекулд хадгалагддаг бөгөөд эсэд тархдаггүй. Митохондрийн бүтцийг зөрчих нь амьсгалын үйл явцыг тасалдуулж, улмаар бие махбодийн эмгэг өөрчлөлтөд хүргэдэг.
Голги аппарат.Голги аппарат (синоним - диктомиом) нь 3-12 хавтгай хэлбэртэй, хаалттай, танк хэмээх давхар мембран дискээр хүрээлэгдсэн, ирмэгээс нь олон тооны бөмбөлгүүд цайруулж өгдөг (300-500). Савны өргөн нь 6-90 А, мембраны зузаан нь 60-70 А байна.
Голги аппарат нь полисахарид, ялангуяа целлюлозыг нэгтгэх, хуримтлуулах, тусгаарлах төв бөгөөд уургийн тархалт, эсийн доторх эсийн тээвэрлэлт, түүнчлэн вакуол болон лизосом үүсэхэд оролцдог. Ургамлын эсэд Голги аппаратын оролцоо, медиан хавтангийн гаднах байдал, эсийн пекто-целлюлозын мембраны өсөлтийг ажиглах боломжтой байв.
Голги аппарат нь эсийн идэвхитэй амьдралд хамгийн их хөгжсөн байдаг. Нас ахих тусам аажмаар атрофийн шинж тэмдэг илэрч, дараа нь алга болдог.
Лизосомууд.Лизосомууд - арай жижиг (диаметр нь 0.5 микрон) бөөрөнхий биетэй. Тэдгээр нь уураг-липоид мембранаар бүрсэн байдаг. Лизосомын агуулга нь уургийн макромолекул, нуклейн хүчил, полисахаридын эсийн доторхи хоол боловсруулах (лизис) функцийг гүйцэтгэдэг олон тооны гидролит ферментүүд юм. Тэдний гол үүрэг бол эсийн протопластын бие даасан хэсгийг хоол боловсруулах (аутофаги - өөрийгөө залгих). Энэ процесс нь фагоцитоз эсвэл пиноцитозын улмаас үргэлжилдэг. Энэ үйл явцын биологийн үүрэг хоёр талтай юм. Нэгдүгээрт, энэ нь хамгаалалтын шинж чанартай, учир нь түр зуурын нөөцийн хомсдол нь эс нь үндсэн уураг болон бусад бодисоос үүдэлтэй амьдралыг дэмждэг бөгөөд хоёрдугаарт илүүдэл буюу элэгдсэн органеллээс (пластид, митохондри гэх мэт) ялгардаг. эс бөгөөс эдгээр ферментүүд бүрэн боловсруулж авах болно.
Үхсэн эсэд лизосомууд устаж, ферментүүд эсэд байдаг бөгөөд түүний бүх агуулга шингэдэг. Үлдсэн зүйл бол пекто-целлюлозын бүрхүүл юм.
Лизосом нь Голги аппаратын үйл ажиллагааны бүтээгдэхүүн бөгөөд үүнээс салсан судас бөгөөд энэ эрхтэн нь хоол боловсруулах фермент хуримтлагддаг.
Сферосом - 0.3-0.4 микрон хэмжээтэй бөөрөнхий уураг-липоид биетэй. Магадгүй бүх зүйлд эдгээр нь Голги аппаратын буюу эндоплазмын торлогийн дериватив юм. Тэдний хэлбэр, хэмжээ нь лизосомтой төстэй байдаг. Сферосом нь хүчил фосфатаза агуулдаг тул лизосомуудтай холбоотой байж болох юм. Зарим зохиогчид сферосом ба лизосомууд бие биентэйгээ тэнцүү боловч хамгийн ихдээ зөвхөн гарал үүсэл, хэлбэртэй байдаг гэж үздэг. Тэдний өөх тосыг нийлэгжүүлэхэд оролцдог гэсэн таамаглал байдаг (А. Фрей-Вислинг).
Рибосомууд - маш жижиг органоид, диаметр нь 250А орчим байдаг ба бараг бөмбөрцөг хэлбэртэй байдаг. Тэдгээрийн зарим нь эндоплазмын торлог бүрхэвчийн гаднах мембран дээр бэхлэгдсэн байдаг бол зарим хэсэг нь цитоплазмын чөлөөт байдалд байдаг. Нэг эс 5 сая хүртэлх рибосом агуулж болно. Рибосомууд нь хлоропластууд ба митохондруудад байдаг ба эдгээр органоидууд болох уургийн нэг хэсэг, тэдгээрийн доторх ферментүүдийг синтезжүүлдэг.
Гол үүрэг нь цөмөөс авсан мэдээллийн дагуу тодорхой уургийн нийлэгжилт юм. Тэдгээрийн найрлага: уураг ба рибосомын рибонуклеины хүчил (РНХ) -ийг ижил хувь хэмжээгээр агуулдаг. Тэдний бүтэц нь рибонуклеотидээс үүссэн жижиг, том дэд хэсгүүд юм.
Микротубулууд.Микротубулууд - эндоплазмын торлог бүрхэвчийн өвөрмөц деривативууд. Олон нүдэнд олддог. Тэдний нэр нь тэдгээрийн хэлбэрийн тухай ярьдаг - зэрэгцээ байрладаг нэг буюу хоёр, дотор нь хөндий бүхий хоолой. 250 А дотор байгаа диаметр. Микротубулуудын хана нь уургийн молекулуудаас бүрддэг. Эсийн хуваагдлын үед спиндний судлууд нь микротубулуудаас үүсдэг.
Гол нь
Ургамлын эс дотор бөөмийг 1831 онд Р. Браун нээсэн. Энэ нь эсийн төвд буюу эсийн мембраны ойролцоо байрладаг боловч бүх талаараа цитоплазмаар хүрээлэгдсэн байдаг. Ихэнх тохиолдолд эсэд нэг цөм байдаг, хэд хэдэн бөөм нь зарим замаг, мөн мөөгөнцрийн эсэд байдаг. Эсийн бус ногоон замаг нь хэдэн зуун цөмтэй байдаг. Бөөгнөрөлгүй лакакуудын олон тооны эсүүд. Бактери ба цэнхэр ногоон замагны эсүүд дотор бөөм байхгүй.
Цөмийн хэлбэр нь бөмбөг эсвэл эллипсийн хэлбэртэй хамгийн ойр байдаг. Эсийн хэлбэр, нас, үйл ажиллагаанаас хамаарна. Морицист эсэд цөм нь том, бөөрөнхий хэлбэртэй бөгөөд эсийн эзэлхүүний 3/4 -ийг эзэлдэг. Эпидермисийн паренхимал эсүүд нь том төв вакуолтой байдаг ба цөм нь лентикуляр хэлбэртэй бөгөөд цитоплазмын хамт эсийн зах руу шилждэг. Энэ бол мэргэшсэн, гэхдээ аль хэдийн хөгширсөн эсийн шинж тэмдэг юм. Цөм дутагдалтай эс нь богино хугацаанд л амьдарч чадна. Шигшүүрийн хоолойн цөмийн эсүүд нь амьд эсүүд боловч урт насалдаггүй. Бусад бүх тохиолдолд цөмгүй эсүүд үхсэн байдаг.
Цөм нь давхар бүрхүүлтэй, дотор нь нүх сүвээр дамждаг
цөм (нуклеоплазм) нь цитоплазмын агууламжтай харьцаж чаддаг. Бөөмийн мембраны мембран нь рибосомоор тоноглогдсон бөгөөд эсийн эндоплазмын торлог бүрхэвчинтэй холбогддог. Нуклеоплазмд нэг буюу хоёр нуклеоли ба хромосом байрладаг. Нуклеоплазм нь Золагийн коллоид систем бөгөөд өтгөрүүлсэн желатинтай төстэй. Оросын биохимийн судлаачдын (Збарский И.Б., бусад.) Мэдээлснээр цөм нь уургийн дөрвөн фракц агуулдаг: энгийн уураг - глобулин 20%, дезоксирибонуклеопротеид - 70%, хүчил уураг - 6%, үлдэгдэл уургууд 4% байна. Тэдгээр нь дараахь цөмийн бүтцэд нутагшсан байдаг: ДНХ-ийн уураг (шүлтлэг уураг), хромосом дахь РНХ уураг (хүчиллэг уураг), зарим хэсэг нь хромосомуудад (элч РНХ синтез хийх явцад), цөмийн мембран дотор байдаг. Глобулинууд нь нуклеоплазмын үндэс болдог. Үлдэгдэл уургууд (шинж чанар нь тодорхойлогдоогүй) нь цөмийн мембран үүсгэдэг.
Гол уургийн дийлэнх хэсэг нь ДНХ дээр суурилсан цогц шүлтлэг уураг, дезоксирибонуклеопротеид юм.
ДНХ молекул.ДНХ молекул - полинуклеотид ба нуклеотидуудаас тогтдог. Нуклеотид нь гурван бүрэлдэхүүн хэсгээс бүрдэнэ: чихрийн молекул (дезоксирибоз), азотын молекул, фосфорын хүчлийн молекул. Деоксирибоз нь азотын суурь гликозидтой, фосфорын хүчилтэй - эфиртэй холбоотой байдаг. ДНХ-д янз бүрийн хослуулан азотын суурь дээр бие биенээсээ ялгаатай нуклеотидын ердөө 4 сорт байдаг. Тэдгээрийн хоёр (аденин ба гуанин) нь пурин азотын нэгдлүүд, цитозин ба тимин пиримидинд хамаардаг. ДНХ молекулууд нэг хавтгайд байрладаггүй боловч хоёр спираль хэлхээнээс тогтдог. хоёр зэрэгцээ гинж нэг нөгөөг нь тойрон эргэлдэж нэг ДНХ молекул үүсгэдэг. Тэд азотын суурийн хооронд устөрөгчийн холбоо үүсгэн бие биетэйгээ холбогддог бөгөөд нэг гинжин хэлхээний пурин суурь нь нөгөөдөх пиримидиний суурьтай холбогддог (Зураг 14). ДНХ молекулын бүтэц, химийг Англи (Крик) ба Америкийн (Ватсон) эрдэмтэд нээж, 1953 онд олон нийтэд танилцуулжээ. Энэ цэг нь молекулын генетикийн хөгжлийн эхлэл гэж тооцогддог. ДНХ-ийн молекулын жин нь 4-8 сая.Нуклеотидын тоо (янз бүрийн хувилбарууд) 100 мянга хүртэл байдаг. ДНХ молекул нь маш тогтвортой бөгөөд түүний тогтвортой байдал нь түүний зузаан нь ижил байдаг - 20А (8А - пиримидин баазын өргөн + 12А - пурин баазын өргөн). Хэрэв цацраг идэвхт фосфорыг биед нэвтрүүлсэн бол шошгыг ДНХ-ээс бусад бүх фосфор агуулсан нэгдлүүдээс олж авах болно (Леви, Сикевиц).
ДНХ молекулууд нь удамшлын тээгч юм, учир нь тэдгээрийн бүтэц нь бие махбодийн шинж чанарыг тодорхойлдог тодорхой уургийн нийлэгжилтийн талаарх мэдээллийг кодчилдог. Өөрчлөлт нь мутагеник хүчин зүйлийн нөлөөн дор (цацраг идэвхт цацраг, хүчтэй, химийн бодис, алкалоид, спирт гэх мэт) нөлөөлж болно.
РНХ молекул.Рибонуклеины хүчил молекул (РНХ) цөөн ДНХ молекулууд. Эдгээр нь нуклеотидын нэг гинж юм. Гурван төрлийн РНХ байдаг: рибосомаль, хамгийн урт, олон тооны гогцоо үүсгэдэг, мэдээлэл (матриц) ба тээвэр, хамгийн богино. Рибосомаль РНХ нь эндоплазмын торлог бүрхэвчийн рибосомуудад нутагшдаг ба эсийн нийт РНХ-ийн 85% -ийг эзэлдэг.
Мэдээллийн РНХ нь түүний бүтэц дэх хошоонгор навчтай адил юм. Түүний хэмжээ нь эс доторх нийт РНХ-ийн 5% байдаг. Энэ нь бөөмд нийлэгждэг. Түүний угсралт нь интерфазын үед хромосомуудад тохиолддог. Үүний гол үүрэг бол уургийн нийлэгжилт явагддаг ДНХ-ээс рибосом руу мэдээлэл дамжуулах явдал юм.
Тээврийн РНХ нь одоо тогтсон байдлаар бүтэц, биологийн үйл ажиллагаатай холбоотой бүхэл бүтэн нэгдэл юм. Бараг тооцооллоор амьд эс бүхэн 40-50 бие даасан РНХ агуулдаг ба зүйлийн ялгаа нь харгалзан үзэгдэх шинж чанараараа асар том юм. (Акад. V. Энгельхардт). Тэдний молекулууд уургийн нийлэгжилтийн эсийн доторх эсийн үйл явцыг тээвэрлэх үйлчилгээ эрхэлдэг тул тэдгээрийг тээвэр гэж нэрлэдэг. Үнэгүй амин хүчлүүдтэй нийлээд тэдгээрийг барьж буй уургийн гинжин хэлхээний рибосомуудад хүргэдэг. Эдгээр нь дунджаар 80 нуклеотидээс бүрдэх хамгийн жижиг РНХ молекул юм. Цитоплазмын матрицад байршуулж, эсийн РНХ-ийн 10 орчим хувийг эзэлдэг
РНХ нь азот агуулсан дөрвөн суурийг агуулдаг боловч ДНХ-ээс ялгаатай нь uracil нь РНХ-д тимины оронд байдаг.
Хромосомын бүтэц.Хромосомыг анх 19-р зууны сүүлээр цитологийн Флеминг ба Страсбургер (1882, 1884) сонгодог бүтээлээр олж нээсэн бөгөөд Оросын эс судлаач И.Д. Чистяков тэднийг 1874 онд олж нээжээ.
Хромосомын үндсэн бүтцийн элемент бол цөм юм. Тэд өөр хэлбэртэй. Эдгээр нь шулуун эсвэл муруй саваа, зууван биет, бөмбөг хэлбэртэй, хэмжээ нь өөр өөр байдаг.
Центромерийн байршлаас хамааран шууд, тэгш, тэгш бус хромосомуудыг ялгаж үздэг. Хромосомын дотоод бүтцийг Зураг дээр үзүүлэв. 15, 16. Дезоксирибонуклеопротеин нь хромосомын мономер гэдгийг тэмдэглэх нь зүйтэй.
Дезоксирибонуклеопротеины хромосомын 90-92% нь үүний 45% нь ДНХ, 55% нь уураг (гистон) юм. РНХ (мэдээлэл) нь хромосомд бага хэмжээгээр агуулагддаг.
Хромосом нь мөн тод томруун бүтэцтэй байдаг - өтгөрүүлсэн хэсгүүд байдаг - 1909 онд буцаж ирсэн дискүүд. генүүд гэж нэрлэдэг. Энэ нэр томъёог Данийн эрдэмтэн Йохансен санал болгов. 1911 онд Америкийн эрдэмтэн Морган генүүд нь удамшлын гол нэгжүүд бөгөөд хромосомуудад шугаман дарааллаар тархдаг болохыг нотолж, улмаар хромосом нь чанарын хувьд өөр өөр хэсгүүдтэй болжээ. 1934 онд Америкийн эрдэмтэн Пинтер хромосомын морфологийн бүтэц, хромосом дахь дискүүд байгаа эсэхийг нотолж, дискүүд нь ДНХ хуримтлагддаг газар юм. Энэ нь организмын нэг буюу өөр шинж чанарыг тодорхойлдог генийн байршлыг (локус) зааж өгсөн хромосомын газрын зураглалыг эхлүүлэх эхлэл болсон юм. Ген гэдэг нь нэг уургийн бүтцийн талаархи мэдээллийг агуулсан ДНХ-ийн давхар спираль бүс юм. Энэ бол нэг уургийн молекулын нийлэгжилтийг тодорхойлдог ДНХ молекулын хэсэг юм. ДНХ нь уургийн нийлэгжилтэд шууд оролцдоггүй. Энэ нь зөвхөн уургийн бүтцийн талаархи мэдээллийг агуулдаг бөгөөд хадгалдаг.
Дараалан байрлах 4 нуклеотидын хэдэн мянгаас бүрдэх ДНХ бүтэц нь удамшлын код юм.
Удамшлын хууль. Уургийн нийлэгжилт.ДНХ-ийн кодын тухай анхны мессежийг 1961 онд Америкийн биохимик Ниренберг Москвад Олон улсын биохимийн конгресст хийсэн. ДНХ-ийн кодын мөн чанар нь дараах байдалтай байна. Амин хүчил бүр гурван зэргэлдээ нуклеотидын (трипл) ДНХ-ийн нэг хэсэгтэй тохирч байна. Жишээлбэл, TT-T (3 тимин агуулсан нуклеотидын гурвалсан хэсэг) нь амин хүчил лизин, триплет А (аденин) - C (цитозин) - A (аденин) - цистеин гэх мэттэй таарч байна. Генийг дараах дарааллаар зохион бүтээсэн нуклеотидын гинжээр төлөөлнө гэж бодъё: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G. Энэ цувралыг гурвалсан хэлбэрээр задалсны дараа бид ямар амин хүчил, ямар дарааллаар нийлэгжсэн уурагт байрлахыг даруй олж тогтооно.
Гурав дахь 4 нуклеотидын боломжтой хослолын тоо 4х64 байна. Эдгээр харьцааг үндэслэн илүүдэл өөр өөр гурвалсан тоо нь бие махбодийн бүтэц, функцийг тодорхойлдог олон тооны уургийн нийлэгжилтийн талаархи мэдээллийг өгөхөд хангалттай байх болно. Уургийн синтезийн хувьд энэхүү мэдээллийн яг хуулбарыг элч РНХ-ийн хэлбэрээр рибосом руу илгээдэг. I-RNA-ээс гадна олон төрлийн тээврийн рибонуклеины хүчил (т-РНХ), рибосом, олон тооны ферментүүдийн молекулууд нь задрал, синтезд оролцдог. 20 амин хүчил тус бүр нь Т-РНХ - молекултай молекултай холбогддог. 20 амин хүчил тус бүр өөрийн Т-РНХ байдаг. Т-РНХ нь амин хүчилээ "таних" чадвартай химийн бүлэгтэй бөгөөд үүнийг боломжтой амин хүчлүүдээс нь сонгон авдаг. Энэ нь тусгай ферментийн тусламжтайгаар тохиолддог. Амин хүчлийг нь мэдээд t-RNA нь түүнтэй нэгдэлд ордог. Р-РНК-ийн гинжин хэлхээний (молекулын) эхэн хэсэгт рибосом хавсарч, би-РНХ-ийн дагуу урагшилж, энэ I-РНХ-ийн нуклеотидын дарааллаар кодлогддог амин хүчлүүд яг хоорондоо холбогддог. Үүний үр дүнд уургийн молекул үүсдэг бөгөөд түүний найрлага нь генийн аль нэгэнд кодлогддог.
Нуклеоли- цөмийн салшгүй бүтцийн хэсэг. Эдгээр нь бөмбөрцөг бие юм. Тэд маш их өөрчлөгдөж, хэлбэр, бүтэц өөрчлөгдөж, гарч, алга болдог. Нэг, хоёр байна. Ургамал бүрийн хувьд тодорхой тооны. Эс нь хуваагдахад бэлдэж, дараа нь дахин гарч ирэх үед бөөм алга болно; тэд рибонуклеины хүчлийн синтезд оролцож байгаа нь илт. Хэрэв рентген эсвэл хэт ягаан туяаны фокусын туяагаар нуколол уствал эсийн хуваагдал дарагдана.
Эсийн амьдрал дахь цөмийн үүрэг. Бөөм нь эсийн хяналтын төв болж үйлчилдэг бөгөөд эсийн үйл ажиллагааг удирдан чиглүүлж, тухайн организмын онцлог шинж чанарыг тодорхойлдог удамшлын тээгч (ген) агуулдаг. Хэрэв бичил мэс заслын аргыг ашиглан түүнийг эсээс гаргаж аваад үүний үр дагаврыг ажиглавал цөмийн үүргийг тодорхойлж болно. Эсийн өсөлтийг зохицуулахад чухал үүрэг гүйцэтгэдэг болохыг нотолсон цуврал туршилтыг Хеммерлинг нэг эсийн ногоон замаг Acetobularia дээр хийв. Энэ далайн ургамал 5 см өндөрт, мөөг шиг харагддаг, "үндэс", "хөл" хэлбэртэй байдаг. Дээрх нь том хэмжээтэй диск хэлбэртэй "малгай" -аар дуусдаг. Энэхүү замагны эс нь эсийн суурь хэсэгт байрладаг нэг цөмтэй байдаг.
Хаммерлинг хөлөө тайрвал доод хэсэг нь үргэлжлүүлэн ажиллаж, үйл ажиллагааны дараа малгай нь бүрэн шинэчлэгддэг болохыг тогтоожээ. Бөөмөөс салаагүй дээд хэсэг хэсэг хугацаанд оршин тогтнодог боловч эцэст нь доод хэсгийг нь сэргээх чадваргүй болж үхдэг. Иймээс ацетобулийн цөм нь өсөлтийн үндэс дэх бодисын солилцооны урвал шаардлагатай байдаг.
Бөөм нь эсийн хана үүсэхийг дэмждэг. Үүнийг замаг Ваучерия, Спирограатай хийсэн туршилтаар дүрсэлж болно. Зүссэн судсан эсүүдээс усыг гаргаж авснаар бид цитоплазмын бөөгнөрлийг нэг, хэд хэдэн бөөм, цөмгүйгээр авах боломжтой. Эхний хоёр тохиолдолд эсийн мембран хэвийн үүссэн. Цөм байхгүй тохиолдолд бүрхүүл үүсээгүй.
И.И.Герасимовын (1890) спирогиатай хийсэн туршилтаар хоёр цөм эсүүд нь хлоропластын урт ба зузааныг хоёр дахин нэмэгдүүлдэг болохыг тогтоожээ. Цөмийн эсэд фотосинтезийн үйл явц үргэлжилж, ассимиляци цардуул үүсдэг боловч түүний гидролизийн үйл явц идэвхжиж, энэ нь зөвхөн цөмийн ДНХ-ийн дагуу рибосомт нийлэгжих гидролит фермент байхгүй байгаатай холбоотой юм. Протопластын цөмгүй амьдрал нь богино, богино байдаг. Туршилтанд I.I. Герасимовын цөмгүй спирогир эсүүд 42 хоног амьдарч байгаад нас баржээ. Цөмийн хамгийн чухал үүргүүдийн нэг нь цитоплазмыг эсийн уургийн нийлэгжилтэд шаардлагатай рибонуклеины хүчилээр хангах явдал юм. Цөмийг эсээс салгах нь цитоплазм дахь РНХ-ийн агууламж аажмаар буурч, доторх уургийн нийлэгжилт удааширдаг.
Цөмийн хамгийн чухал үүрэг бол шинж чанаруудыг эсээс эс рүү, организмаас организм руу шилжүүлэх явдал бөгөөд үүнийг цөмийн задрал болон эсийн үйл явцад гүйцэтгэдэг.
Эсийн хуваагдал.Эсүүд хуваагдах замаар үрждэг. Энэ тохиолдолд эх эсийн адил хромосомоор бүрхэгдсэн удамшлын материалын ижил багц бүхий нэг эсээс хоёр охин эс үүсдэг. Соматик эсэд хромосомыг хоёр буюу хоёуланг нь төлөөлдөг бөгөөд үүнд генийн хромосом гэж нэрлэгддэг бөгөөд үүнд аллелийн генүүд (эсрэг талын тэмдэгтүүдийг зөөгч, жишээ нь, цагаан, улаан вандуйны дэлбээ гэх мэт), хоёр эцэг эхийн хосын тэмдгийг агуулдаг. Үүнтэй холбоотойгоор ургамлын биен дэх соматик эсүүдэд үргэлж 2p гэсэн тэмдэг бүхий давхар хромос байдаг. Хромосом нь тодорхой шинж чанартай байдаг. Хромосомын тоо хэмжээ, чанар нь зүйл бүрийн онцлог шинж юм. Тиймээс гүзээлзгэнэ эсэд диплоид багц хромос 14, (2н), алимны мод 34, Иерусалим артишок 102 г.м байна.
Митоз (караокинез) - соматик эсийн хуваагдлыг анх Е.Русов (1872), И.Д.Чистяков (1874) нар тайлбарласан байдаг. Үүний мөн чанар нь ижил эсийн хромосом бүхий хоёр охин эс нь эх эсээс хуваагдах замаар үүсдэгт \u200b\u200bоршино. Микроавтобографийн арга нь хамгийн урт бөгөөд хамгийн хэцүү нь интерфаза буюу "амрах" цөмийн үе юм гэдгийг тогтоожээ. энэ хугацаанд цөмийн материалын хэмжээ хоёр дахин нэмэгдэж байна. Интерфаза нь гурван үе шатанд хуваагдана.
Q1 - presynthetic (түүний үргэлжлэх хугацаа 4-6 цаг);
S - нийлэг (10-20 цаг);
Q2 - постинтетик (2-5 цаг).
Q1 үе шатанд ДНХ-ийн хувилах бэлтгэл ажил хийгдэж байна. Мөн S үе шатанд ДНХ багасч, эс нь ДНХ-ний нийлүүлэлтийг хоёр дахин нэмэгдүүлдэг. Q2 үе шатанд митозыг өдөөхөд шаардлагатай фермент, бүтэц бий болдог. Тиймээс интерфазад хромосом дахь ДНХ молекулууд хоёр ижил хэлхээнд хуваагддаг бөгөөд мэдээллийн РНХ нь тэдний матриц дээр угсардаг. Сүүлийнх нь цитоплазм дахь тодорхой уургийн бүтцийн талаархи мэдээллийг авч үздэг бөгөөд ДНХ-ийн судалт бүр нь молекулынхаа хагасыг алга болгодог. Хоёр дахин нэмэгдэх (багасгах) үйл явцад ДНХ-ийн өвөрмөц шинж чанар илэрдэг бөгөөд энэ нь ДНХ-ийн өөрийгөө зөв үржих чадвараас бүрддэг. Үүссэн охин ДНХ молекулыг эцэг эх молекулын яг хуулбараар автоматаар авдаг, учир нь хуулбарлах үед хүрээлэн буй орчноос нэмэлт (АТ; Г-Ц; г.м.) баазууд тал бүр дээр нэгддэг.
Митозын хуваагдлын үед хоёр дахин томрох хромосомууд мэдэгдэхүйц болдог. Метафазад тэд бүгд экваторын бүсэд, нэг эгнээнд байрладаг. Бөөгнөрлийн судлууд үүсдэг (бие биетэйгээ холбогдож буй микротюбуудаас). Бөөм ба цөмийн бүрхүүл алга болдог. Зузаан хромосомыг хоёр охин хромосом болгон уртын дагуу хуваана. Энэ бол митозын мөн чанар юм. Энэ нь охин эсийн хоорондох ДНХ-ийн давхар молекулын үнэн зөв хуваарилалтыг өгдөг. Энэ нь ДНХ-д шифрлэгдсэн удамшлын мэдээллийг дамжуулах боломжийг олгодог.
Анафазад охины хромосомууд эсрэг туйл руу шилжиж эхэлдэг. Төв хэсэгт эсийн мембраны эхний хэсгүүд (фрагмобласт) гарч ирдэг.
Телофазад бөөмүүд эсийн эсүүдэд үүсдэг. Эх эсийн агууламж (органелл) нь үүссэн охины эсүүдийн хооронд тараагддаг. Эсийн мембран нь бүрэн үүссэн байдаг. Энэ нь цитокинезийг бүрэн төгс болгодог (Зураг 17).
Meiosis - бууруулах хэлтэс үүнийг өнгөрсөн зууны 90-ээд оны үед В.И.Беляев нээсэн ба тайлбарласан байдаг. Хэсэгчлэх мөн чанар нь 2н (давхар, диплоид) хромос агуулсан соматик эсээс дөрвөн хаплоид эс, хагас хромосом бүхий "n" үсэгтэй байдаг. Энэ төрлийн хуваагдал нь нарийн төвөгтэй бөгөөд хоёр үе шатаас бүрдэнэ. Эхнийх нь хромосомын бууралт юм. Давхардсан хромосомууд экваторын бүсэд хосоороо байрладаг (хоёр ижил төстэй гомолог хромосомууд). Энэ үед хромозоор дамжих (наалдац), хөндлөн гарах (кроссовер) үүсч болох ба үүний үр дүнд хромозын цэгүүдийн солилцоо үүсч болно. Үүний үр дүнд эцэг эхийн хромосомын генүүдийн нэг хэсэг нь эхийн хромосомын хэсэг болж, эсрэгээр үүсдэг. Үүний үр дүнд хоёулангийнх нь хромосомын гадаад байдал өөрчлөгдөхгүй боловч чанарын найрлага нь өөр болж хувирдаг. Аав, эхийн удамшил нь дахин хуваарилагдаж, холилдсон байдаг.
Миозын анафазын үед гомологийн хромосомууд нь туйл руу салж, хэсэг хугацааны дараа богино байдаг (судлууд алга болж, харин шинэ бөөмүүдийн хоорондох септум үүсдэггүй), митозын үйл явц эхэлдэг - бүх хромосомууд нэг хавтгайд байрладаг метафаза, тэдгээрийн урт хэсэг нь хуваагддаг. охин хромосом дээр. Анафазын митозын тусламжтайгаар гинжний тусламжтайгаар туйлууд хоорондоо хуваагддаг бөгөөд тэнд дөрвөн бөөм үүсдэг ба үүний үр дүнд дөрвөн гаплоид эсүүд үүсдэг. Хөгжих явцад зарим эд эсийн эсэд тодорхой хүчин зүйлийн нөлөөн дор бүрэн бус митоз үүсч, тэдгээр нь туйлд хуваагддаггүйн улмаас бөөм дэх хромосомын тоо хоёр дахин нэмэгддэг. Байгалийн эсвэл хиймэл шинж чанарыг ийм зөрчлийн үр дүнд тетраплоид ба полиплоидууд үүсдэг. Миозын тусламжтайгаар үр хөврөлийн эсүүд үүсдэг - гаметууд, түүнчлэн спор, ургамлын бэлгийн болон асексуал нөхөн үржихүйн элементүүд (Зураг 18).
Амитоз нь цөмийн шууд задрал юм. Амитозын үед хуваагдмал холбоос үүсдэггүй бөгөөд митозын нэгэн адил цөмийн бүрхүүл ялзардаггүй. Өмнө нь амитозыг хуваах үндсэн хэлбэр гэж үздэг байсан. Энэ нь бие махбодийн доройтолтой холбоотой болохыг одоо тогтоожээ. Энэ нь цөмийг илүү төвөгтэй хуваагдлын хялбаршуулсан хувилбар юм. Амитоз нь бөөм, эндосперм, булцууны паренхим, навчны дэлбээ гэх мэт эс, эдэд тохиолддог.
Митохондриа - Энэ нь спираль, бөөрөнхий, сунасан эсвэл салаалсан органелл юм.
Митохондрийн тухай ойлголтыг анх Бенда 1897 онд дэвшүүлсэн. Митохондрийг үр тариа, мөхлөг эсвэл судалтай хэлбэрээр фазын тодосгогч ба хөндлөнгийн микроскоп ашиглан амьд эсүүдэд илрүүлж болно. Эдгээр нь хөдлөх, бие биетэйгээ нийлэх, хуваалцах чадвартай хөдөлгөөнт бүтэц юм. Хөнгөн микроскопийн дор үхсэн эсүүдэд тусгай аргаар будсан бол митохондриа нь цитоплазмд ялимгүй тархсан эсвэл зарим тодорхой бүсэд төвлөрсөн жижиг үр тариа (мөхлөг) мэт харагддаг.
Хүчилтөрөгч байгаа үед глюкоз ба өөх тосыг устгасны үр дүнд митохондрияд энерги үүсч, органик бодис ус, нүүрстөрөгчийн давхар исэл болж хувирдаг. Ийм маягаар амьтны организм нь амьдралын шаардлагатай энергийг авдаг. Эрчим хүч нь аденозин трифосфат (ATP), илүү нарийвчлалтайгаар түүний макроэлектрик бондод хуримтлагддаг. Митохондрийн үйл ажиллагаа нь органик нэгдлүүдийн исэлдэлт, тэдгээрийн задралын үеэр ялгарах энергийг ATP молекулын нийлэгжилтэд ашиглахтай нягт холбоотой байдаг. Тиймээс митохондрийг эсийн энергийн станц буюу эсийн амьсгалын замын органелл гэж нэрлэдэг. ATP нь энерги нийлүүлэгчээр ажилладаг бөгөөд энерги баялаг терминал фосфатын бүлгийн нэг хэсгийг өөр молекул руу шилжүүлж ADP болж хувирдаг.
Хувьслын үед митохондри нь эртний эсийн биед симбиотик болсон прокариот бичил биетнүүд байсан гэж үздэг. Дараа нь тэд дэлхийн агаар мандалд хүчилтөрөгчийн агууламж нэмэгдсэнтэй холбоотойгоор зайлшгүй шаардлагатай болсон. Нэг талаас, митохондриа нь эсийн илүүдэл хортой хүчилтөрөгчийг зайлуулж, нөгөө талаас энерги өгдөг.
Митохондри байхгүй бол эс нь хүчилтөрөгчийг эрчим хүчээр хангадаг бодис болгон ашиглах чадваргүй бөгөөд зөвхөн энергийн хэрэгцээгээ анаэробийн процессоор хангах боломжтой юм. Ийнхүү хүчилтөрөгч нь хор, харин хор нь эсийн хувьд амин дэм бөгөөд илүүдэл хүчилтөрөгч нь түүний дутагдалтай адил хортой юм.
Митохондриа нь хэлбэрээ өөрчилж, эсийн хэрэгцээ хамгийн их байгаа тэр хэсгүүдэд шилжиж болно. Тиймээс кардиомиоцитын үед митохондри нь миофибрилийн ойролцоо, бөөрний гуурсан хоолойн эсийн эсийн ойролцоо байрладаг гэх мэт. Эс нь мянга хүртэлх митохондри агуулдаг бөгөөд тэдгээрийн тоо нь эсийн идэвхжилээс хамаардаг.
Митохондри нь дунджаар 0.5 ... 3 микрон хэмжээтэй байдаг. Хэмжээнээс хамааран жижиг, дунд, том, аварга том митохондри ялгаж авдаг (тэдгээр нь салаалсан сүлжээ үүсгэдэг - митохондрийн торлог бүрхэвч). Митохондрийн хэмжээ, тоо нь эсийн үйл ажиллагаа, түүний эрчим хүчний хэрэглээтэй нягт холбоотой байдаг. Эдгээр нь маш их хувьсах чадвартай бөгөөд эсийн идэвхжил, хүчилтөрөгчийн агууламж, дааврын нөлөөллөөс шалтгаалан хавдаж, өөрчлөгдөж, кристаагийн тоо, бүтэц өөрчлөгдөж, тоо, хэлбэр, хэмжээ, ферментийн идэвхээс хамаардаг.
Митохондрийн дийлэнх нягтрал, тэдгээрийн дотоод гадаргуугийн хөгжлийн түвшин болон бусад үзүүлэлтүүд нь эсийн эрчим хүчний хэрэгцээнээс хамаардаг. Лимфоцитуудад цөөн тооны митохондри байдаг ба элэгний эсүүдэд 2 ... 3 мянга байдаг.
Митохондриа нь матриц, дотоод мембран, перимитохондрийн орон зай, гадна талын мембранаас бүрдэнэ. Гаднах митохондрийн мембран нь органеллийг гиалоплазмаас тусгаарладаг. Ихэвчлэн энэ нь гөлгөр контуртай бөгөөд хаалттай байдаг тул мембран уут юм.
Дотор талаас гаднах мембраныг 10 ... 20 нм өргөнтэй перимитохондрийн орон зайгаар тусгаарладаг. Дотоод митохондрийн мембран нь дотоод митохондрийн дотоод агуулгыг хязгаарладаг - матриц. Дотор мембран нь хавтгай уулархаг хэлбэртэй, эсвэл cristae хэлбэртэй байдаг митохондриас дотогшоо олон тооны дэгдэлт үүсгэдэг.
Хөнгөн хэлбэртэй бол cristae нь ялтсууд (трабекуляр) ба гуурс хэлбэртэй (зүсэлт дээр мультивикуляр) хэлбэртэй байж болох бөгөөд тэдгээр нь митохондрийн хувьд уртааш буюу хөндлөн чиглэлд чиглэгддэг.
Митохондри тус бүр матрицаар дүүрэн байдаг бөгөөд энэ нь электрон микроографикт хүрээлэн буй цитоплазмаас илүү нягт харагддаг. Митохондрийн матриц нь янз бүрийн электрон нягтрал бүхий нэг төрлийн (нэгэн төрлийн), заримдаа нарийн ширхэгтэй байдаг. Үүний дотор 2 ... 3 нм зузаантай нимгэн судлууд, 15 ... 20 нм хэмжээтэй ширхэгүүд илэрдэг. Матрицын судлууд нь ДНХ молекул, жижиг мөхлөгүүд нь митохондриал рибосомууд юм. Матриц нь фермент, нэг судалтай, цикл ДНХ, митохондриал рибосом, олон тооны Ca 2+ ион агуулдаг.
Митохондрийн уургийн нийлэгжилтийн бие даасан системийг гистон агуулаагүй ДНХ молекулаар төлөөлдөг. ДНХ нь богино, цагираг хэлбэртэй (цикл) бөгөөд 37 ген агуулдаг. Цөмийн ДНХ-ээс ялгаатай нь түүний дотор кодлогдоогүй нуклеотидын дараалал бараг байдаггүй. Бүтэц, зохион байгуулалтын онцлог нь нянгийн эсийн ДНХ-тэй митохондрийн ДНХ-ийг нэгтгэдэг. Митохондрийн ДНХ дээр янз бүрийн хэлбэрийн РНХ молекулуудын нийлэгжилт үүсдэг: мэдээлэл, дамжуулалт (тээвэрлэлт) ба рибосомал. Митохондрийн мэдээллийн РНХ-ийг үрэлтэд оруулдаггүй (мэдээллийн ачааллыг авч чадахгүй зүсэлт хийх). Митохондрийн ДНХ-ийн молекулуудын жижиг хэмжээ нь бүх митохондрийн уургийн нийлэгжилтийг тодорхойлж чадахгүй. Ихэнх митохондрийн уургууд нь эсийн цөмийн генетикийн хяналтанд байдаг бөгөөд цитоплазмд нийлэгждэг тул митохондрийн ДНХ нь маш сул илэрхийлэгддэг бөгөөд исэлдүүлэгч фосфорлантын гинжин ферментийн зөвхөн нэг хэсгийг бүрдүүлж чаддаг. Митохондрийн ДНХ нь мембран дотор байрладаг арав гаруй уургийг кодлодог бөгөөд бие махбодийн функциональ уургийн цогцолборыг зөв нэгтгэх үүрэгтэй бүтэцтэй уураг юм. Тээврийн функцийг гүйцэтгэдэг уураг мөн нийлэгждэг. Уургийн нийлэгжилтийн ийм систем нь митохондрийн бүх функцийг хангаж чаддаггүй тул митохондрийн бие даасан байдал хязгаарлагдмал, харьцангуй байдаг.
Хөхтөн амьтдад бордох үед митохондри нь зөвхөн өндөгөөр дамждаг бөгөөд эр бэлгийн эс нь шинэ организмд нуклейн ДНХ нэвтрүүлдэг.
Митохондрийн матрицад цитоплазмын рибосомуудаас ялгаатай рибосомууд үүсдэг. Тэд цөмөөр кодлогдоогүй олон тооны митохондрийн уургийн нийлэгжилтэнд оролцдог. Митохондрийн рибосомууд нь тунадасны тоо 60-тай байдаг (цитоплазмаас тунадасны тоо 80-тай ялгаатай). Тунадасны тоо нь центрифугал болон хэт төвлөрсөн үед хуримтлагдсан хэмжээ юм. Бүтэцийн хувьд митохондриал рибосом нь прокариот организмын рибосомтой ойролцоо боловч жижиг хэмжээтэй бөгөөд тодорхой антибиотик (хлорамфеникол, тетрациклин гэх мэт) -д мэдрэмтгий байдаг.
Дотоод митохондрийн мембран нь бодисыг зөөвөрлөх чадвар өндөртэй байдаг. Исэлдүүлэгч фосфоржуулалтын гинжин холбоо, электрон зөөгч уургууд, ATP, ADP, пируват болон бусад тээврийн системүүд хоорондоо ойрхон байрлах түүний дотоод гадаргуу дээр бэхлэгддэг.Дотоод мембран дээр ферментийн нягт зохицуулалт хийсний үр дүнд биохимийн процессын өндөр уялдаа холбоо (харилцан хамаарал) нэмэгддэг. каталитик үйл явцын үр ашиг.
Электрон микроскопоор матрицын люмен руу цухуйсан мөөгний хэсгүүд илэрдэг. Тэд ATP-нийлэг (ADP-ээс ATP үүсгэдэг) үйл ажиллагаатай байдаг. Электрон тээвэр нь дөрвөн том ферментийн цогцолбор (цитохром) агуулсан дотоод мембран дотор байрлах амьсгалын замын гинжин хэлхээний дагуу явагддаг. Амьсгалын замаар дамжуулж электронууд устөрөгчийн ионыг матрицаас перимитохондрийн орон зайд шахдаг бөгөөд энэ нь протон градиент (насос) үүсэхийг баталгаажуулдаг. Энэхүү градиентийн энерги (бодисын концентраци болон мембраны потенциал үүсэх ялгаа) нь АТФ нийлэгжих, метаболит ба органик бус ионуудыг тээвэрлэхэд ашиглагддаг. Дотор мембран дахь уураг агуулсан уураг нь органик фосфат, ATP, ADP, амин хүчил, өөх тосны хүчил, три- болон дикарбон хүчлийг дамжуулдаг.
Гитрофилийн уургийн олон сувагтай тул гаднах митохондрийн мембран нь бага молекул жинтэй бодисуудад илүү нэвчилттэй байдаг. Тусгай рецепторын цогцолборууд гаднах мембран дээр байрладаг бөгөөд тэдгээрийн тусламжтайгаар матрицаас уураг нь перимитохондрийн орон зайд дамждаг.
Химийн найрлага, шинж чанарын хувьд гаднах мембран нь бусад эсийн эсийн мембран, плазмолемматай ойролцоо байдаг. Энэ нь өөх тосыг метаболизмд оруулдаг, амин хүчил, амин исэлдүүлэгч болох хувиргалтыг идэвхжүүлдэг (катализлуулдаг) фермент агуулдаг. Хэрэв гаднах мембраны ферментүүд идэвхтэй хэвээр байвал энэ нь митохондрийн үйл ажиллагааны аюулгүй байдлын үзүүлэлт юм.
Митохондрийн хоёр бие даасан дэд хэсэг байдаг. Перимитохондрийн орон зай буюу гаднах митохондрийн танхим (гаднах дэд тасалгаа) нь гиалоплазмын уургийн цогцолбор нэвтрэн орсноос үүсдэг тул дотоод дэд тасалгаа (митохондрийн матриц) нь митохондрийн ДНХ-ийн синтетик үйл ажиллагааны улмаас хэсэгчлэн үүсдэг. Дотоод дэд тасалгаа (матриц) нь ДНХ, РНХ, рибосом агуулдаг. Энэ нь гиалоплазмтай харьцуулахад өндөр 2 Ca ион агуулдаг. Гаднах дэд тасагт устөрөгчийн ионууд хуримтлагддаг. Гадны болон дотоод дэд хэсгүүдийн ферментатив идэвхжил, уургийн найрлага нь маш өөр өөр байдаг. Дотоод дэд тасалгаа нь гаднахаас илүү электрон нягтралтай байдаг.
Тодорхой митохондрийн тэмдэглэгээ нь цитохромын оксидаза ба сукцинат дегидрогеназын ферментүүд бөгөөд үүнийг илрүүлэх нь митохондрия дахь энергийн процессын тоон үзүүлэлтүүдийг тодорхойлох боломжийг олгодог.
Митохондрийн үндсэн үүрэг - ATP синтез. Эхэндээ элсэн чихэр (глюкоз) нь гиалоплазм дотор сүүн ба пирувийн хүчил (пируват) -ийг бага хэмжээний АТФ синтезээр устгадаг. Нэг глюкозын молекулын гликолизийн үр дүнд хоёр ATP молекул ашиглагддаг бөгөөд дөрөв нь үүсдэг. Тиймээс зөвхөн хоёр ATP молекул нь эерэг тэнцвэрийг бүрдүүлдэг. Эдгээр үйл явц нь хүчилтөрөгчгүй (анаэробик гликолиз) явагддаг.
Эрчим хүчний үйлдвэрлэлийн дараагийн бүх үе шатууд нь их хэмжээний ATP-ийн нийлэгжилтийг хангадаг аэроб исэлдэлтийн процесст явагддаг. Энэ тохиолдолд органик бодис CO 2 болон устай хамт устдаг. Исэлдэлт нь протоныг хүлээн авагчид шилжүүлэхтэй хамт явагддаг. Эдгээр урвалыг митохондрийн матрицад байрладаг трикарбоксилийн хүчлийн олон тооны ферментүүд ашиглан гүйцэтгэдэг.
Электрон дамжуулалтын систем ба холбогдох фосфоржилтын ADP (исэлдүүлэгч фосфоризаци) нь булчирхайн мембранд байрладаг. Энэ тохиолдолд нэг электрон хүлээн авагч уургаас электроныг нөгөөд шилжүүлэх бөгөөд эцэст нь хүчилтөрөгчтэй холбож, улмаар ус үүсдэг. Үүний зэрэгцээ электрон шилжүүлэх гинжин хэлхээнд ийм исэлдэлт гарах үед ялгарч буй энергийн нэг хэсэг нь ADP фосфоржилтийн үед макроэлектрик бонд хэлбэрээр хадгалагддаг бөгөөд энэ нь олон тооны ATP молекул - эсийн эсийн энерги үүсгэдэг. Исэлдэлтийн фосфоржуулалтын процесс нь митохондриал cristae-ийн мембран дээр исэлдэлтийн гинжин уураг, энд байрладаг ADP ATP синтетаза фосфоризацийн ферментийг ашиглан явагддаг. Исэлдүүлэгч фосфоржуулалтын үр дүнд нэг глюкозын молекулаас 36 ATP молекул үүсдэг.
Митохондрийн зарим даавар, бодисын хувьд тусгай (хамаарал бүхий) рецепторууд байдаг. Триодиотиронин нь ихэвчлэн митохондрийн синтетик идэвхийг хурдасгадаг. Триодиотиронины үл тэвчих исэлдүүлэгч фосфоржуулалтын гинжний интерлейкин-1 ба өндөр концентраци нь митохондрийн хавдар үүсгэдэг бөгөөд энэ нь дулааны энерги үүсэх дагалддаг.
Шинэ митохондри нь хуваагдмал, нягтрал эсвэл нахиалах замаар үүсдэг. Сүүлчийн тохиолдолд protomitochondria үүсч, хэмжээ нь аажмаар нэмэгддэг.
Protomitochondria бол гаднах болон дотор талын мембран бүхий жижиг органелл юм. Дотор мембран нь хөгжөөгүй cristae агуулаагүй байна. Органелла нь аэробик фосфоржилтын түвшин бага байдаг. Бодисжилт үүссэнээр митохондрийн агуулгыг хоёр шинэ нэлээд том органеллуудын хооронд тараана. Нөхөн үржихүйн аливаа аргыг ашиглан шинээр үүссэн митохондри тус бүр өөрийн геномтой байдаг.
Хөгшин митохондри нь автолизосом үүсгэхийн тулд аутолиз (лизосомыг ашиглан эсийн өөрөө боловсруулж) устгадаг. Автолизосомоос үлдэгдэл бие үүсдэг. Бүрэн хоол боловсруулснаар бага молекул жинтэй органик бодисоос бүрдэх үлдэгдэл бие махбодид эксоцитоз ялгардаг. Бүрэн боловсруулалгүйгээр митохондрийн үлдэгдэл эсэд хуримтлагдаж, нипофусцин бүхий ширхэгтэй бие буюу мөхлөг хэлбэрээр үүсдэг. Уусдаггүй кальцийн давс нь митохондрийн хэсэгт хуримтлагдаж, талстууд үүсдэг. Митохондрийн доройтлын бүтээгдэхүүний хуримтлал нь эсийн дистрофи үүсгэдэг.
Митохондрийн бүтэц, үйл ажиллагаа нь нэлээд төвөгтэй асуудал юм. Органелл байх нь бараг бүх цөмийн организмд хамааралтай байдаг - эдгээр нь бараг бүх амьтад, зарим ургамал, мөөгөнцөр болох автотроф (фотосинтез хийх чадвартай ургамал) болон гетеротрофийн хувьд юм.
Митохондрийн үндсэн зорилго нь органик бодисын исэлдэлт, энэ процессын үр дүнд ялгарсан энергийг дараагийн хэрэглээ юм. Энэ шалтгааны улмаас органеллууд бас хоёрдахь (албан бус) нэртэй байдаг - эсийн энергийн станцууд. Тэднийг заримдаа "катаболизмын пластид" гэж нэрлэдэг.
Митохондриа гэж юу вэ
Энэ нэр томъёо нь Грек гаралтай. Орчуулсан нь энэ үг нь "утас" (митос), "үр" (chondrion) гэсэн утгатай. Митохондри нь эсийн хэвийн үйл ажиллагаанд чухал ач холбогдолтой байнгын организмоид бөгөөд бүхэл бүтэн организм оршин тогтнох боломжийг олгодог.

"Станцууд" нь тодорхой дотоод бүтэцтэй байдаг ба энэ нь митохондрийн үйл ажиллагааны байдлаас хамаарч өөр өөр байдаг. Тэдний хэлбэр нь хоёр хэлбэртэй байж болно - зууван эсвэл гонзгой. Сүүлийнх нь ихэвчлэн мөчир хэлбэртэй байдаг. Нэг эсийн органеллуудын тоо 150-1500 хооронд хэлбэлздэг.
Онцгой тохиолдол бол үр хөврөлийн эсүүд юм. Спермэд ганцхан спираль органелл байдаг бол эмэгтэй хүний \u200b\u200bгаметад хэдэн зуун мянган илүү митохондри агуулагддаг. Эсийн дотор органеллууд нэг газарт тогтдоггүй боловч цитоплазмын дагуу хөдөлж, бие биентэйгээ нийлж чаддаг. Тэдний хэмжээ 0.5 микрон, урт нь 60 микрон хүрч чаддаг бол хамгийн бага хэмжээ нь 7 микрон болно.

Нэг "цахилгаан станц" -ын хэмжээг тодорхойлох нь амаргүй ажил биш. Баримт нь электрон микроскопоор шалгаж үзэхэд органеллээс зөвхөн хэсэг нь зүсмэл рүү ордог. Энэ нь спираль митохондри нь бие даасан, бие даасан бүтцэд авах хэд хэдэн хэсэгтэй байдаг.
Зөвхөн гурван хэмжээст дүрс нь эсийн яг нарийн бүтцийг олж, 2-5 бие даасан органоид эсвэл митохондрийн нарийн төвөгтэй хэлбэрийн тухай ярьж байгаа эсэхийг ойлгох боломжийг олгоно.
Бүтцийн онцлог
Митохондрийн мембран нь гадаад ба дотоод гэсэн хоёр давхаргаас бүрдэнэ. Сүүлийнх нь навч хэлбэртэй, гуурсан хэлбэртэй янз бүрийн гацалт, атираа орно.
Мембран бүр нь тусгай химийн найрлагатай, тодорхой хэмжээний фермент, тодорхой зорилготой байдаг. Дотор талаас гаднах бүрхүүл нь 10-20 нм зузаантай дундын хоорондын зайгаар тусгаарлагддаг.
Зураг дээр органеллийн бүтэц нь маш тодорхой харагдаж байна.

Митохондрийн бүтэц
Бүтцийн бүдүүвчийг харахдаа дараахь зүйлийг хийж болно. Митохондрия доторх наалдамхай орон зайг матриц гэж нэрлэдэг. Түүний найрлага нь түүнд шаардлагатай химийн процессын урсгалын таатай орчинг бүрдүүлдэг. Энэ нь урвал ба биохимийн процессыг дэмжих микроскопийн мөхлөгт агуулдаг (жишээлбэл, тэд гликоген ион болон бусад бодисыг хуримтлуулдаг).
Матрицад ДНХ, коэнзим, рибосом, т-РНХ, органик бус ионууд орно. ATP синтаз ба цитохромууд нь бүрхүүлийн дотоод давхаргын гадаргуу дээр байрладаг. Ферментүүд нь Кребсийн мөчлөг (CTK), исэлдүүлэгч фосфоржилт гэх мэт үйл явцад хувь нэмэр оруулдаг.
Тиймээс, органоидын гол ажлыг матриц ба бүрхүүлийн дотор талыг хоёуланг нь гүйцэтгэдэг.
Митохондрийн үйл ажиллагаа
"Цахилгаан станцууд" -ын зорилгыг хоёр үндсэн ажилаар тодорхойлж болно.
- эрчим хүчний үйлдвэрлэл: исэлдэлтийн процессууд нь тэдгээрийн дотор ATP молекулуудыг ялгаруулах замаар явагддаг;
- генетикийн мэдээллийг хадгалах;
- гормон, амин хүчил болон бусад бүтцийг нийлүүлэх ажилд оролцдог.
Исэлдэлт, энерги үйлдвэрлэх үйл явц хэд хэдэн үе шаттайгаар явагддаг.


ATP синтезийн схемийн зураг
Үүнийг тэмдэглэх нь зүйтэй юм. Кребсийн цикл (нимбэгийн хүчлийн цикл) -ийн үр дүнд ATP молекул үүсэхгүй, молекулууд исэлдэн, нүүрстөрөгчийн давхар исэл ялгардаг. Энэ бол гликолиз ба электрон тээврийн гинжин хэлхээний хоорондын завсрын алхам юм.
Хүснэгт "Митохондрийн функц ба бүтэц"

Нэг эс дэх митохондрийн тоог юу тодорхойлдог вэ
Эрх мэдлийн нөөц шаардлагатай байгаа эсийн ойролцоо байгаа олон тооны органоид хуримтлагддаг. Ялангуяа миофибрилийн хэсгүүдэд олон тооны органелл цуглуулдаг бөгөөд энэ нь тэдний агшилтыг баталгаажуулдаг булчингийн эсүүдийн нэг хэсэг юм.

Эрэгтэй үр хөврөлийн эсүүдэд бүтэц нь flagellum тэнхлэгийн эргэн тойронд нутагшдаг - гампийн сүүлний тогтмол хөдөлгөөнөөс болж ATP-ийн хэрэгцээ үүсдэг гэж үздэг. Хөдөлгөөнд зориулагдсан тусгай cilia ашигладаг protozoa дахь митохондрияны зохион байгуулалт нь органеллууд нь яг ижил байдлаар мембран дор хуримтлагддаг мэт харагдаж байна.
Мэдрэлийн эсийн тухайд мэдрэлийн системийн дохио дамждаг синапсийн ойролцоо митохондрийн нутагшуулалт ажиглагддаг. Уураг нийлэгжүүлдэг эсүүдэд органеллууд эргастоплазмын бүсэд хуримтлагддаг - энэ үйл явцыг хангадаг энерги өгдөг.
Митохондрийг хэн илрүүлэв
Эсийн бүтэц 1897-1898 онд C. Брэндийн ачаар нэрээ олж авав. Отто Вагбург 1920 онд эсийн амьсгалын замын митохондриатай холбож байгааг баталж чадсан.
Дүгнэлт
Митохондриа нь амьд эсийн хамгийн чухал бүрэлдэхүүн хэсэг бөгөөд ATP молекулыг үүсгэдэг энергийн станцын үүрэг гүйцэтгэдэг бөгөөд ингэснээр эсийн амин чухал үйл явцыг хангадаг.
Митохондрийн ажил нь органик нэгдлүүдийн исэлдэлт дээр суурилдаг тул эрчим хүчний потенциал үүсдэг.
Гаднах мембран
Дотор талын мембран
Матриц матриц, кристаБайна. энэ нь гөлгөр контуртай, тайралт, атираа үүсгэхгүй. Энэ нь бүх эсийн мембраны талбайн 7 орчим хувийг эзэлдэг. Түүний зузаан нь 7 нм орчим бөгөөд энэ нь цитоплазмын бусад мембрануудтай холбоогүй бөгөөд өөрөө хаалттай байдаг тул мембран уут юм. Гаднах мембран нь дотроосоо тусгаарлагдсан байдаг хоорондын зай ойролцоогоор 10-20 нм өргөн. Дотоод мембран (ойролцоогоор 7 нм зузаантай) нь митохондрийн дотоод агууламжийг хязгаарлаж,
түүний матриц буюу митоплазм. Митохондрийн дотоод мембраны онцлог шинж чанар нь тэдний митохондри дотор олон тооны дэгдэлт үүсгэх чадвар юм. Ийм дэгдэлт нь ихэнхдээ хавтгай уулархаг хэлбэртэй байдаг. Криста дахь мембраны хоорондох зай 10-20 нм орчим байна. Ихэнхдээ cristae нь хуруу хэлбэртэй процессыг салгаж, хэлбэржүүлж, нугалж, тодорхой чиглэлтэй байдаггүй. Протозо, нэг эсийн замаг, өндөр ургамал, амьтдын зарим эсэд дотоод мембраны гаднах байдал хуруу шилэн (tubular cristae) шиг харагддаг.
Митохондри матриц нь нарийн ширхэгтэй, нэгэн төрлийн бүтэцтэй, заримдаа бөмбөлгөнд цуглуулсан нимгэн судалтай (ойролцоогоор 2-3 нм), 15-20 нм орчим ширхэг байдаг. Одоо митохондрийн матрицын судлууд нь митохондрийн нуклеоидын найрлага дахь ДНХ молекулууд, жижиг мөхлөгүүд нь митохондриал рибосомууд юм.
Митохондрийн үйл ажиллагаа
1. Митохондриад ATP нийлэгжилт явагдана (Исэлдүүлэгч фосфоржилтийг үзнэ үү)
Intermembrane pH ~ 4, матрицын рН ~ 8 | уургийн агууламж м: 67% нь матриц, 21% нь уурхайн нээлт, 6% нь уурхайн дотор, 6% нь уурхай хоорондын
Хуурах - нэг митохондрийн систем
гаднах m-na: порин-нүх нь 5 кД хүртэл дамжуулалт хийх боломжийг олгодог. | дотоод m-na: ионуудад зориулсан кардиолипин-үл нэвтрэх м-ху |
олон улсын үйлдвэрлэл: ферментийн бүлгүүд фосфорилатын нуклеотид ба нуклеотидын сахар
Дотроо:
матриц: бодисын солилцооны ферментүүд - липидийн исэлдэлт, нүүрс усны исэлдэлт, трикарбоксилийн хүчлийн мөчлөг, Кребсийн цикл
Бактериас гаралтай: amoeba Pelomyxa palustris нь эукариотуудаас нэгдмэл байдаггүй, аэробик бактери бүхий симбиозид амьдардаг | өөрийн ДНХ | OX төстэй бактери
Митохондрийн ДНХ
Миохондрийн хуваагдал
хуулбарласан
интерфазад | хуулбар нь S-фаз | -тай холбоогүй болно cl мөчлөгийн үед митозууд нэг удаа хоёр хуваагдаж, нягтрал үүсгэдэг бөгөөд дотор нь нягтрал үүсдэг m-not | ~ 16.5 кб | бөгж 2 rRNA 22 tRNA ба 13 уураг кодчилдог |
уургийн тээвэрлэлт: сигнал пептид | амфилийн муруй | митохондрийн таних рецептор |
Исэлдүүлэгч фосфоржилт
Электрон дамжуулалтын хэлхээ
ATP синтез
элэгний эсүүдэд м ~ митохондрийн хуваагдал үүсэх замаар 20 хоног амьдардаг
16569 bp \u003d 13 уураг, 22tRNA, 2pRNA | гөлгөр гаднах m-na (поринууд - 10 кДа хүртэлх уургийн нэвчилт) (атираат) уураг (75% уураг): тээврийн зөөгч уураг, f-та, амьсгалын гинжин бүрэлдэхүүн хэсэг ба ATP синтаз, кардиолипин) матриц ( f-tami цитрат циклээр баяжуулсан) inter 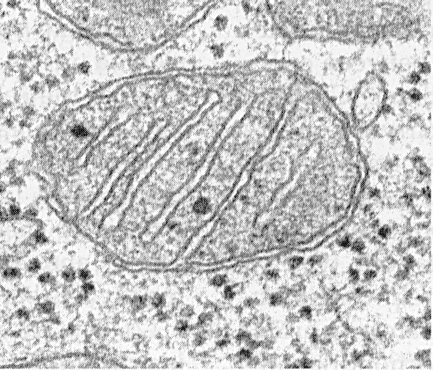
Митохондриа буюу хондриосомууд (грек хэлнээс. Mitos - утас, chondrion - үр тариа, soma - бие) нь protozoa, ургамал, амьтдын цитоплазмд байдаг мөхлөгт эсвэл судалтай органелл юм. Митохондриа нь амьд эсүүдэд ажиглагдаж болох тул нэлээд өндөр нягтралтай байдаг. Амьд эсийн доторх митохондри нь бие биентэйгээ хөдөлж, хөдөлж, нэгдэж чаддаг.
Янз бүрийн зүйлийн хувьд митохондрийн хэмжээ маш их, мөн хэлбэр нь өөрчлөгддөг (Зураг 199). Гэсэн хэдий ч ихэнх эсүүдэд эдгээр бүтцийн зузаан харьцангуй тогтмол байдаг (ойролцоогоор 0.5 мкм), урт нь хэлбэлзэж, судалтай хэлбэрээр 7-60 мкм хүртэл хүрдэг.
Митохондрийн хэмжээ, тоог судлах нь тийм ч энгийн зүйл биш юм. Энэ нь хэт нимгэн хэсгүүд дээр харагдах митохондрийн хэмжээ, тоо хэмжээ нь бодит байдалтай тохирохгүй байгаатай холбоотой юм.
Ердийн тооцоогоор элэгний эсэд 200 орчим митохондри байдаг. Энэ нь нийт цитоплазмын 20 гаруй хувь, эс доторх нийт уургийн 30-35 хувийг эзэлдэг. Элэгний эсийн бүх митохондрийн гадаргуугийн талбай нь түүний плазмын мембраны гадаргуугаас 4-5 дахин том байдаг. Ихэнх митохондри нь ооцит (300,000 орчим) ба аварга том амеба эмх замбараагүй эмх замбараагүй байдалд (500,000 хүртэл) байдаг.
Ногоон ургамлын эсэд митохондрийн тоо амьтны эсийнхээс бага байдаг тул хлоропластууд үйл ажиллагааныхаа нэг хэсгийг гүйцэтгэдэг.
Эсийн доторх митохондрийн нутагшуулалт өөр өөр байдаг. Ихэвчлэн митохондрия нь цитоплазмын тэр хэсгүүдийн ойролцоо хуримтлагддаг бөгөөд үүнд митохондри үүссэн ATP шаардлагатай байдаг. Тиймээс, араг ясны булчинд митохондри нь миофибрилийн ойролцоо байрладаг. Спермийн хувьд митохондри нь flagellum тэнхлэгийн эргэн тойронд спираль хэлбэртэй байдаг; энэ нь эр бэлгийн эсийн сүүлийг хөдөлгөхөд ATP ашиглах шаардлагатай байгаатай холбоотой юм. Үүнтэй адилаар protozoa болон cilia-ээр тоноглогдсон бусад эсүүдэд митохондриа нь эсийн мембран дор cilia-ийн суурь дээр байрладаг бөгөөд энэ нь ATP-ийг ажиллуулахыг шаарддаг. Мэдрэлийн эсийн axons-д мэдрэлийн импульс дамжуулах үйл явц явагддаг синапсийн ойролцоо митохондри байрладаг. Их хэмжээний уураг синтез хийдэг нууц эсүүдэд митохондри нь эргастоплазмын бүсүүдтэй нягт холбоотой байдаг; магадгүй рибосом дээр амин хүчил, уургийн нийлэгжилтийг идэвхжүүлэхийн тулд ATP нийлүүлдэг байх.
Митохондрийн хэт авиан бүтэц.
Митохондриа нь хэмжээ, хэлбэрээс үл хамааран бүх нийтийн бүтэцтэй байдаг бөгөөд тэдгээрийн хэт авианы бүтэц жигд байдаг. Митохондриа нь хоёр мембранаар хязгаарлагддаг (Зураг 205). Гаднах митохондрийн мембран нь үүнийг гиалоплазмаас тусгаарладаг, жигд контуртай, цухуйж, атираа үүсгэдэггүй, зузаан нь 7 нм орчим байдаг. Энэ нь бүх эсийн мембраны талбайн 7 орчим хувийг эзэлдэг. Мембран нь цитоплазмын бусад мембрануудтай холбогдоогүй, өөрөө хаалттай, мембран уут юм. Дотор талаас гаднах мембран нь ойролцоогоор 10-20 нм өргөнтэй дундын хоорондын зайгаар тусгаарлагддаг. Дотоод мембран (ойролцоогоор 7 нм зузаантай) нь митохондри, түүний матриц буюу митоплазмын дотоод агууламжийг хязгаарладаг. Дотоод митохондрийн мембран нь митохондрийн дотор олон тооны томрол үүсгэдэг. Ийм дэгдэлт нь ихэнхдээ хавтгай уулархаг хэлбэртэй байдаг.
Элэгний эс доторх дотоод митохондрийн мембраны нийт гадаргуу нь бүх эсийн мембраны гадаргуугийн гуравны нэг орчим байдаг. Зүрхний булчингийн эсийн митохондри нь элэгний митохондриас гурав дахин их байдаг бөгөөд энэ нь өөр өөр эсийн митохондрийн функциональ ачааллын ялгааг харуулдаг. Криста дахь мембраны хоорондох зай 10-20 нм орчим байна.
Митохондриал cristae нь дотоод мембран дээрээс гарч матриц руу чиглэсэн байдаг тул митохондрийн хөндийг бүрэн бөглөрдөггүй бөгөөд матрицыг дүүргэх тасралтгүй байдлыг зөрчдөггүй.
Митохондрийн урт тэнхлэгтэй холбоотой кристаагийн чиглэл нь өөр өөр эсүүдэд өөр өөр байдаг. Зүг чиг баримжаа нь перпендикуляр (элэг, бөөр) cristae байж болно; зүрхний булчинд cristae-ийн уртааш зохицуулалт ажиглагддаг. Криста нь хуруу хэлбэртэй процессыг салгаж, хэлбэржүүлж, нугалж, тодорхой чиглэлтэй байдаггүй (Зураг 208). Протозо, нэг эсийн замаг, өндөр ургамал, амьтдын зарим эсэд дотоод мембраны гаднах байдал хуруу шилэн (tubular cristae) шиг харагддаг.
Митохондри матриц нь нарийн ширхэгтэй нэг төрлийн бүтэцтэй бөгөөд ДНХ молекулыг бөмбөг (2-3 нм орчим) орчим цуглуулсан нимгэн судалтай, митохондриал рибосомууд нь ойролцоогоор 15-20 нм хэмжээтэй мөхлөг хэлбэрээр илэрдэг. Матриц дахь магни, кальцийн давсны ордууд нь том хэмжээтэй (20-40 нм) өтгөн мөхлөг үүсгэдэг.
Митохондрийн үйл ажиллагаа.
Митохондриа нь органик субстратыг исэлдүүлэх, ADP-ийн фосфоржуулах үйл явцын үр дүнд үүсдэг ATP-ийн синтезийг явуулдаг.
Нүүрс ус исэлдүүлэх эхний үе шатыг anaerobic исэлдүүлэх гэж нэрлэдэг гликолиз ба гиалоплазмд тохиолддог бөгөөд хүчилтөрөгчийн оролцоог шаарддаггүй. Анаэробын энерги үйлдвэрлэх явцад исэлдүүлэх субстрат нь гексоз, юуны өмнө глюкоз юм; зарим бактериуд пентоз, өөх тос эсвэл амин хүчлийг исэлдүүлэх замаар энерги гаргаж авах чадвартай байдаг.
Глюкозын хувьд C, H, O атомуудын хоорондын бондонд агуулагдах боломжит энергийн хэмжээ нь нэг мэнд ойролцоогоор 680 ккал байдаг (жишээ нь, 180 г глюкоз).
Амьд эсийн дотор ийм их хэмжээний энерги нь үе шаттайгаар явагддаг бөгөөд олон тооны исэлдүүлэгч ферментүүдээр удирддаг бөгөөд химийн холболтын энерги нь шаталтын үед халуунд шилжихтэй холбоогүй боловч ATP молекул дахь макроэнергетик холбоо болж хувирдаг бөгөөд эдгээрээс ялгарсан энерги ашиглан нийлэгддэг. ADP ба фосфат.
Гликолизийн үр дүнд үүссэн триозууд, үндсэндээ пирувийн хүчил нь митохондрийн цаашдын исэлдэлтэд оролцдог. Энэ тохиолдолд бүх химийн бондоос ялгарах энерги ашиглагддаг бөгөөд энэ нь CO 2-ыг ялгаруулж хүчилтөрөгчийн хэрэглээ болон их хэмжээний АТФ нийлэгжихэд хүргэдэг. Эдгээр процессууд нь трикарбоксилын хүчлийн исэлдэлтийн мөчлөг, электрон дамжуулалтын амьсгалын гинжин холбоо, ADP фосфоризаци, эсийн "түлш" ба ATP молекулуудын нийлэгжилт үүсдэг (Зураг 209).
Трикарбоксилын хүчлийн мөчлөгт (Кребсийн цикл эсвэл нимбэгийн хүчлийн цикл) гликолизын үр дүнд үүссэн пируват нь эхлээд CO 2 молекулыг алдаж ацетатад (хоёр нүүрстөрөгчийн нэгдэл), коэнзим А-тай нэгддэг. Дараа нь ацетил коэнзим А нь оксалацетаттай (а, дөрвөн) зургаан нүүрстөрөгчийн цитрат (нимбэгийн хүчил) үүсгэдэг. Дараа нь энэ зургаан нүүрстөрөгчийн нэгдлийг дөрвөн нүүрстөрөгчийн оксалацетаттай исэлдүүлэх цикл байдаг бөгөөд энэ нь ацетил коэнзим А-д дахин холбогддог ба дараа нь цикл давтагдана. Энэ исэлдэлтийн үед хоёр CO 2 молекул ялгарч, исэлдэлтийн явцад ялгарсан электронууд нь хүлээн авагч коэнзим молекулууд руу шилждэг (NAD-никотинамид аденины динуклеотид) бөгөөд энэ нь цаашлаад электрон дамжуулах хэлхээнд ордог. Тиймээс, трикарбоксилын хүчлүүдийн циклд өөрөө АТФ нийлэгжилт байдаггүй, харин молекулын исэлдэлт, электроныг хүлээн авагчид шилжүүлэх, CO 2-ын ялгаруулалт байдаг. Дээр дурьдсан бүх үйл явдлууд митохондри нь тэдний матрицад явагддаг.
Анхны субстратыг исэлдүүлэх нь CO 2 ба ус ялгаруулахад хүргэдэг боловч шаталтын явцад дулааны энерги ялгардаггүй боловч ATP молекулууд үүсдэг. Тэдгээр нь исэлдэлттэй шууд холбоогүй өөр нэг уургийн бүлгээр нийлэгждэг. Митохондрийн дотоод мембран дээр матриц руу хардаг мембраны гадаргуу дээр том уургийн цогцолбор, фермент, ATP синтетазууд байрладаг. Электрон микроскопоор тэдгээр нь матриц руу хардаг мембраны гадаргууг бүхэлд нь чиглүүлж, "мөөг" гэж нэрлэгддэг бие махбодь хэлбэрээр харагддаг. Бие нь хөл, толгойтой, диаметр нь 8-9 нм юм. Иймээс исэлдэлтийн гинжин хэлхээний болон ATP синтезийн ферментийн аль алины ферментүүд дотоод митохондрийн мембрануудад байршдаг (Зураг 201б).
Амьсгалын гинж нь митохондри дахь энергийг хувиргах гол систем юм. Энд амьсгалын замын гинжин хэлхээний элементүүдийг дараалан исэлдүүлэх, нөхөн сэргээх үйл явц явагддаг бөгөөд үүний үр дүнд энерги нь жижиг хэсгүүдэд ялгардаг. Энэхүү энергийн ачаар гинжин хэлхээний гурван цэг дээр АТФ үүсч, ADP ба фосфат үүсдэг. Иймээс исэлдэлт (электрон дамжуулалт) нь фосфоржилттай холбоотой байдаг (ADP + Fn → ATP, жишээлбэл исэлдэлтийн фосфоржилт үүсдэг.
Электрон тээвэрлэх явцад ялгарч буй энерги нь мембран дээр протон градиент хэлбэрээр хадгалагддаг. Митохондрийн мембран дахь электрон дамжуулалтын үед амьсгалын замын хэлхээний цогцолбор бүр протон (эерэг цэнэг) -ийг мембранаар дамжин протоны (эерэг цэнэг) матрицаас мембрана хоорондын зай руу шилжихэд мембран дээрх боломжит ялгаа үүсэхэд хүргэдэг: мембран хоорондын орон зайд эерэг цэнэг давамгайлж, сөрөг - митохондрийн матрицын талаас. Потенциал ялгаа (220 мВ) -т хүрэхэд ATP синтетазын уургийн цогцолбор протоныг матриц руу буцааж аажмаар энергийн нэг хэлбэрийг нөгөө хэлбэрт шилжүүлж эхэлдэг бөгөөд энэ нь ADP болон органик бус фосфатаас ATP үүсгэдэг. Энэ бол исэлдүүлэх процессыг нийлэг, ADP-ийн фосфоржуулалттай хослуулах явдал юм. Субстратууд исэлдэж байхад протонууд дотоод митохондрийн мембранаар шахагдаж байх үед үүнтэй холбоотой ATP синтез үүсдэг. исэлдүүлэгч фосфоржилт явагддаг.
Эдгээр хоёр процессыг салгаж болно. Энэ тохиолдолд субстратыг исэлдүүлэхтэй адил электрон дамжуулалт үргэлжилж байгаа боловч ATP синтез хийгддэггүй. Энэ тохиолдолд исэлдэлтийн үед ялгарч буй энерги нь дулааны энерги болж хувирдаг.
Нян дахь исэлдүүлэгч фосфоржилт.
Фосфоржуулалтыг исэлдүүлэх чадвартай прокариот эсүүдэд трикарбоксилын хүчлийн мөчлөгийн элементүүд цитоплазмд шууд байршдаг бөгөөд амьсгалын замын гинжин хэлхээ ба фосфоризаци ферментүүд нь эсийн мембрантай, цитоплазмын дотор цухуйж, мезосом гэж нэрлэгддэг. Ийм бактерийн мезозом нь зөвхөн аэробик амьсгалын үйл явцтай холбоотой байж болох төдийгүй зарим зүйл эсийн хуваагдал, шинэ эсэд ДНХ тархах, эсийн хананд үүсэх гэх мэт үйл явцад оролцдог болохыг тэмдэглэх нь зүйтэй.
Зарим бактерийн мезосом дахь плазмын мембран дээр исэлдэлт ба ATP синтезийн хоёулангийн нийлмэл процесс явагддаг. Эукариот эсийн митохондриятай төстэй бөмбөрцөг хэсгүүд нь нянгийн плазмын мембраны фракцуудаас электрон микроскопоор олдсон. Ийнхүү исэлдүүлэх фосфоржуулах чадвартай бактерийн эсүүдэд сийвэнгийн мембран нь эукариот эсийн дотоод митохондрийн мембрантай төстэй үүрэг гүйцэтгэдэг.
Митохондрийн тоо нэмэгдэх.
Митохондри нь эсийн хуваагдал эсвэл эсийн функциональ ачааллыг нэмэгдүүлэх замаар тэдний тоог нэмэгдүүлэх боломжтой. Митохондриа байнга шинэчлэгдэж байдаг. Жишээлбэл, элэгний хувьд митохондрийн дундаж наслалт 10 хоног байна.
Өмнөх митохондрийн өсөлт, хуваагдлаар митохондрийн тоо нэмэгдэж байна. Энэ таамаглалыг анх биобластууд гэсэн нэр томъёогоор митохондрийг тодорхойлсон Алтман (1893) илэрхийлжээ. Бактерийн хуваагдлын хоёртын арга хэлбэртэй байдаг vivo хуваагдмал, урт митохондрийг богино хэсгүүдэд хувааж үзэх боломжтой.
Эд эсийн соёлын амьд эсүүд дэх митохондриын төлөв байдлыг судлахдаа хуваах замаар митохондриийн бодит өсөлтийг тогтоов. Эсийн мөчлөгийн үеэр митохондриа хэд хэдэн микрон хүртэл ургадаг ба дараа нь фрагмент жижиг биетүүдэд хуваагддаг.
Митохондриа нь бие биетэйгээ нийлж, зарчмын дагуу үржиж болно: митохондриас митохондриа.
Митохондрийн аутопродукт.
Хоёр мембраны органеллууд нь авто нөхөн үржихүйн бүрэн системтэй байдаг. Митохондри ба пластидын үед ДНХ-ийн мэдээлэл, дамжуулалт ба рибосомын РНХ ба рибосомууд нийлэгжиж митохондри болон пластидын уургийн нийлэгжилтийг явуулдаг. Гэсэн хэдий ч эдгээр системүүд бие даасан боловч тэдгээрийн боломж хязгаарлагдмал байдаг.
Митохондрия дахь ДНХ нь гистонгүй цикл молекул бөгөөд бактерийн хромосомтой төстэй байдаг. Тэдний хэмжээ нь 7 микрон, 16-19 мянган нуклеотидын хос ДНХ нь амьтны митохондрийн нэг цикл молекулд ордог. Хүний биед митохондрийн ДНХ нь 16.5 мянган n.p. агуулдаг бөгөөд энэ нь бүрэн тайлагджээ. Янз бүрийн обьектуудын митохондраль ДНХ нь нэгэн төрлийн шинж чанартай, тэдгээрийн ялгаа нь зөвхөн дотогшоо, хуулбарлагдаагүй газруудын хэмжээнээс хамаардаг болохыг тогтоов. Бүх митохондрийн ДНХ нь бүлэг, кластер хэлбэрээр цуглуулсан олон хуулбар юм. Тиймээс харх элэгний нэг митохондрияд 1-ээс 50 хүртэлх мөчлөгт ДНХ-ийн молекул агуулагдаж болно. Нэг эс тутамд митохондрийн ДНХ-ийн нийт хэмжээ ойролцоогоор нэг хувь байдаг. Митохондрийн ДНХ-ийн синтез нь цөм дэх ДНХ-ийн синтезтэй холбоогүй болно.
Бактерийн нэгэн адил митохондрал ДНХ нь тусдаа бүсэд - нуклеоид, түүний хэмжээ 0.4 микрон диаметртэй байдаг. Урт митохондрийн хувьд 1-ээс 10 хүртэлх нуклеоид байж болно. Урт митохондрийг хуваахдаа нуклеоид агуулсан хэсгийг салгана (бактерийн хоёртын хуваагдалтай төстэй). Хувь хүний \u200b\u200bмитохондрийн нуклеоид дахь ДНХ-ийн хэмжээ нь эсийн төрлөөс хамаарч 10 дахин хэлбэлздэг.
Зарим соёлын хувьд митохондрийн 6-аас 60% -д байдаг эсэд нуклеоид байдаггүй бөгөөд эдгээр органеллүүдийн хуваагдал нь нуклеоидуудын тархалттай бус хуваагдалтай илүү холбоотой байдагтай холбон тайлбарлаж болно.
Өмнө дурьдсанчлан, митохондриа нь хоёулаа хуваагдаж, бие биентэйгээ нийлж чаддаг. Митохондриа бие биетэйгээ нэгдэх үед тэдгээрийн дотоод бүрэлдэхүүн хэсгүүдийн солилцоо үүсч болно.
Митохондриа ба цитоплазмын рРНХ ба рибосомууд эрс ялгаатай болохыг онцлон тэмдэглэх нь зүйтэй. Хэрэв 80-аад оны рибосомууд цитоплазмд байрладаг бол ургамлын эсийн митохондриал рибосом нь 70-аад оны рибосомуудад багтдаг (30 ба 50-ийн дэд хэсгүүдээс бүрддэг, прокариот эсийн онцлог шинжтэй 16, 23-р РНХ агуулдаг), жижиг рибосом нь амьтны эсийн митохондрияд (50 орчим) байдаг.
Митохондрийн рибосомын РНХ нь митохондрийн ДНХ дээр нийлэгждэг. Рибосом дээрх митоплазмд уургийн нийлэгжилт ордог. Бактери дахь уургийн нийлэгжилтийг дарангуйлдаг антибиотик хлорамфениколын нөлөөгөөр цитоплазмын рибосомын нийлэгжилтээс ялгаатай нь энэ нь зогсдог.
Митохондрийн геном дээр 22 тээврийн РНХ нийлэгждэг. Митохондрийн синтетик системийн гурвалсан код нь гиалоплазмд ашигладаг кодоос өөр юм. Уураг нийлүүлэхэд шаардлагатай бүх бүрэлдэхүүн хэсэг байгаа хэдий ч ДНХ-ийн жижиг митохондрийн молекулууд бүх митохондрийн уураг, зөвхөн тэдгээрийн цөөн хэсгийг кодчилдоггүй. Тиймээс ДНХ нь 15 мянган н.п. хэмжээтэй байна. нийт молекул жинтэй 6x10 5 орчим уураг кодлох боломжтой. Үүний зэрэгцээ амьсгалын замын бүрэн бүрэлдэхүүн хэсгийн митохондрийн уургийн уургийн нийт молекулын жин ойролцоогоор 2х10 6-ийн утгад хүрдэг.
Фосфоржуулсан уурагнаас гадна митохондриа нь трикарбоксилын хүчлийн фермент, ДНХ, РНХ синтезийн фермент, амин хүчил идэвхжүүлэх фермент болон бусад уурагуудаас гадна эдгээр олон тооны уураг, рРНХ, тРНХ-ийг кодлохын тулд генетикийн мэдээллийг агуулдаг болохыг бид харж байна. богино митохондрийн ДНХ молекул дутагдалтай байдаг. Хүний митохондрийн ДНХ-ийн нуклеотидын дарааллыг задлахад энэ нь зөвхөн 2 рибосомын РНХ, 22 дамжуулагч РНХ, нийт 13 өөр полипептидийн гинжийг кодлодог болохыг харуулсан.
Ихэнх митохондрийн уураг эсийн эсээс генетикийн хяналтанд байдаг ба митохондрийн гаднах синтез болох нь батлагдсан. Ихэнх митохондрийн уураг цитозол дахь рибосом дээр нийлэгждэг. Эдгээр уургууд нь гаднах митохондрийн мембран дээрх рецепторууд хүлээн зөвшөөрдөг тусгай дохионы дараалалтай байдаг. Эдгээр уургуудыг тэдгээрт оруулж болно (пероксисомын мембраны аналогийг үз), дараа нь дотоод мембран руу шилжинэ. Энэхүү дамжуулалт нь гаднах болон дотор талын мембраны холбоо барих цэгүүдэд, ийм тээвэрлэлтийг тэмдэглэдэг. Ихэнх митохондрийн липидүүд нь цитоплазмд нийлэгждэг.
Энэ бүхэн нь митохондрийн эндосимбиотик гарал үүслийг илтгэнэ, митохондри нь эукариот эсийн хамт симбиозтой байдаг бактери зэрэг организмууд юм.
Чондриом.
Нэг эсийн бүх митохондрияны нэгдлийг chondrioma гэж нэрлэдэг. Энэ нь эсийн төрлөөс хамаарч өөр өөр байж болно. Олон эсийн дотор chondrioma нь цитоплазмын дагуу жигд байрласан эсвэл ATP-ийн эрчимтэй зарцуулалт хийдэг газарт бүлэглэсэн олон тооны митохондриас бүрддэг. Эдгээр хоёр тохиолдолд митохондри нь дангаараа ажилладаг бөгөөд тэдгээрийн хамтын ажиллагаа, магадгүй цитоплазмын зарим дохиогоор зохицуулагддаг. Жижиг тархсан митохондрияны оронд нэг аварга том салаалсан митохондриа эсэд байрладаг бол өөр өөр хэлбэрийн хондриома байдаг.
Ийм митохондри нь нэг эсийн ногоон замаг (жишээлбэл, Chlorella) -д байдаг. Тэдгээр нь нарийн төвөгтэй митохондрийн сүлжээ буюу митохондриал торлог бүрхэвч (Reticulum miyochondriale) үүсгэдэг. Химио-осмотик онолын дагуу гадаад, дотоод мембрануудтай нийлсэн ийм аварга том горхи бүхий митохондрийн бүтцийг бий болгох биологийн утга нь цитоплазмын хаана ч очдог ийм салаалсан митохондрийн дотоод мембраны гадаргуу дээр аль ч цэг дээр ATP синтез үүсч болно гэсэн үг юм. Үүнд шаардлагатай байгаа.
Аливаа цэг дээр аварга том салаалсан митохондри үүссэн тохиолдолд ATP-ийн нийлэгжилтийг эхлүүлэхэд хангалттай потенци нь дотоод мембран дээр хуримтлагдах боломжтой. Эдгээр байрлалаас харахад митохондрийн ретикулум нь цахилгаан дамжуулагч, ийм системийн алслагдсан цэгүүдийг холбосон кабель юм. Митохондрийн торлог бүрхэвч нь зөвхөн хлорелла гэх мэт жижиг хөдөлгөөнт эсүүдэд төдийгүй том бүтэцтэй хэсгүүдэд, жишээлбэл, араг ясны булчин дахь миофибрилуудад маш их хэрэгтэй байв.
Араг ясны булчин нь булчингийн утаснаас бүрддэг, олон цөм агуулсан симпластууд байдаг. Ийм булчингийн утаснуудын урт нь 40 микрон, 0,1 микрон зузаантай байдаг - энэ бол маш олон тооны миофибрилүүдийг агуулдаг маш том бүтэц бөгөөд бүгд нэгэн зэрэг синхрончлогддог. Бэрхшээлийн хувьд агшилтын нэг хэсэг болох миофибрилыг агшилтын нэгж бүрт z-дискний түвшинд өгдөг миофибрилд хүргэдэг. Араг ясны булчингийн хэт авианы хэсгүүдэд электрон микроскопоор саркомерийн ойролцоо байрлах митохондрийн олон тооны жижиг хэсгүүдийг харуулдаг. Булчингийн митохондри нь жижиг бөмбөлөг, эсвэл саваа биш, харин архагнидын бүтэц бөгөөд тэдгээр нь процесс бөгөөд урт зайд, заримдаа булчингийн эсийн бүх диаметрээр дамждаг.
Үүний зэрэгцээ митохондрийн салаа нь булчингийн эс дэх миофибрил бүрийг хүрээлж, булчингийн агшилтанд шаардлагатай ATP-ээр хангадаг. Тиймээс z-дискний хавтгайд митохондри нь ердийн митохондрийн торлог бүрхэвчийг төлөөлдөг. Митохондриал торлог бүрхэвчийн ийм давхарга буюу давхрыг саркомер тус бүрт хоёр удаа давтаж, булчингийн эс бүхэлдээ митохондриал торлог бүрхэвчийн мянга мянган хөндлөн “давхар” давхаргатай байдаг. Миофибриллын дагуух "шалны хооронд" эдгээр митохондрийн давхаргуудыг холбосон судалтай митохондри байдаг. Тиймээс булчингийн эсийн бүхэл бүтэн хэсгийг дамжуулж митохондрийн торлог бүрхэвчийн гурван хэмжээст зургийг бүтээв.
Митохондриал торлог бүрхүүлийн мөчрүүд ба судалтай уртасгасан митохондрийн хэсгүүдийн хооронд тусгай intermitochondrial нэгдлүүд эсвэл контактууд байдаг (MMC). Тэдгээр нь митохондритай холбоо барьж буй гаднах митохондрийн мембрануудаар үүсгэгддэг бөгөөд энэ бүсэд байрлах мембрана хоорондын зай ба мембранууд электрон нягтрал ихэссэн байдаг. Эдгээр тусгай формацаар дамжуулан хөрш зэргэлдээ митохондри ба митохондрийн ретикулумуудын функциональ нэгдэл нь нэгдмэл, нэгдлийн энергийн системд явагддаг. Булчингийн эс дэх бүх миофибрилууд бүхэл бүтэн уртын дагуу синхроноор багасдаг тул ийм нарийн төвөгтэй машины аль ч хэсэгт ATP нийлүүлэх нь синхрон байдлаар явагдах ёстой бөгөөд энэ нь маш олон тооны салаалсан митохондри дамжуулагчууд хоорондоо холбогдсон тохиолдолд л тохиолддог. харилцагчдыг ашиглаж.
Митохондрийн эрчим хүчний холбоонд бие биетэйгээ харилцан холбоотой харилцан үйлчлэл (MMK) үүссэн нь кардиомиоцит, зүрхний булчингийн эсүүд дээр боломжтой байсан.
Зүрхний булчингийн эсүүдийн хондриома нь салаалсан бүтэц үүсгэдэггүй боловч миофибрилуудын хооронд тусгай захиалгагүйгээр байрладаг олон жижиг сунасан митохондри хэлбэрээр илэрдэг. Гэсэн хэдий ч бүх хөрш зэргэлдээ митохондри нь араг ясны булчинд байдаг ижил төрлийн митохондрийн контактуудын тусламжтайгаар бие биетэйгээ нийлдэг, зөвхөн тэдний тоо маш их байдаг: дунджаар митохондрийг нэг хэлхээнд холбодог митохондрияд 2-3 MMC байдаг. гинж (Streptio mitochondriale) нь тусдаа митохондри юм.
Зүрхний эсийн зайлшгүй бүтэц болох intermitochondrial холбоо (MMC) нь бүх сээр нуруутан амьтдын ховдол ба тосгуурын аль алиных нь кардиомиоцитууд болох хөхтөн амьтад, шувууд, хэвлээр явагчид, хоёр нутагтан, ястай загаснаас олдсон нь тодорхой болов. Түүнээс гадна зарим шавьж, нялцгай биетний зүрхний эсэд MMC олдсон (гэхдээ цөөн тоогоор).
Кардиомиоцит дахь MMC-ийн хэмжээ нь зүрхний үйл ажиллагааны ачааллаас хамааран өөр өөр байдаг. Амьтны бие махбодийн идэвхжил нэмэгдэхийн хэрээр MMC-ийн тоо нэмэгдэж, эсрэгээр зүрхний булчинд ачаалал буурах үед MMC-ийн тоо огцом буурдаг.