As the internal structures of mitochondria are called. The structure and functions of mitochondria and plastids
Mitochondria
In the cells of animal tissues, mitochondria were found in 1882, and in plants only in 1904 (in the anthers of water lilies). It was possible to establish biological functions after separation and purification of the fraction by fractional centrifugation. They contain 70% protein and about 30% lipids, a small amount of RNA and DNA, vitamins A, B 6, B 12, K, E, folic and pantothenic acids, riboflavin, various enzymes. Mitochondria have a double membrane, the outer isolates the organelle from the cytoplasm, and the inner forms outgrowths of crista. The entire space between the membranes is filled with a matrix (Fig. 13).
The main function of mitochondria is participation in cellular respiration. The role of mitochondria in respiration was established in 1950-1951. A complex enzyme system of the Krebs cycle is concentrated on the outer membranes. During the oxidation of respiration substrates, energy is released that is immediately accumulated in the formed ADP molecules, mainly ATP, during the oxidative phosphorylation that occurs in cristae. The energy stored in macroergic compounds is used in the future to meet all the needs of the cell.
The formation of mitochondria in the cell occurs continuously from the microbodies, more often their occurrence is associated with the differentiation of the cell membrane structures. They can be restored in the cell by dividing and budding them. Mitochondria are not durable, their life expectancy is 5-10 days.
Mitochondria are “power” cell stations. They concentrate energy, which is stored in the "accumulators" of energy - ATP molecules, and not dissipated in the cell. Violation of the structure of mitochondria leads to disruption of the respiratory process and, as a result, to the pathology of the body.
Golgi apparatus.Golgi apparatus (a synonym - dictiosomes) is a stack of 3-12 flattened, closed, surrounded by a double membrane discs called tanks, from the edges of which numerous bubbles are unfastened (300-500). The width of the tanks is 6-90 A, the thickness of the membranes is 60-70 A.
The Golgi apparatus is a center for the synthesis, accumulation and isolation of polysaccharides, in particular cellulose, and is involved in the distribution and intracellular transport of proteins, as well as in the formation of vacuoles and lysosomes. In the plant cell, it was possible to trace the participation of the Golgi apparatus in the appearance of the median plate and the growth of the cell pecto-cellulose membrane.
The Golgi apparatus is most developed during the period of active cell life. With its aging, it gradually atrophies, and then disappears.
Lysosomes.Lysosomes - rather small (about 0.5 microns in diameter) rounded bodies. They are coated with a protein-lipoid membrane. The contents of lysosomes are numerous hydrolytic enzymes that perform the function of intracellular digestion (lysis) of protein macromolecules, nucleic acids, polysaccharides. Their main function is the digestion of individual sections of the cell protoplast (autophagy - self-devouring). This process proceeds due to phagocytosis or pinocytosis. The biological role of this process is twofold. Firstly, it’s protective, because with a temporary shortage of spare products, the cell supports life due to constitutional proteins and other substances, and secondly, there is a release from excess or worn organelles (plastids, mitochondria, etc.) The lysosome membrane prevents the release of enzymes into the cytoplasm otherwise, it would be completely digested by these enzymes.
In a dead cell, lysosomes are destroyed, enzymes are in the cell and all its contents are digested. All that remains is the pecto-cellulose shell.
Lysosomes are products of the activity of the Golgi apparatus, vesicles detached from it, in which this organoid accumulated digesting enzymes.
Spherosomes - rounded protein-lipoid bodies 0.3-0.4 microns. In all likelihood, they are derivatives of the Golgi apparatus or endoplasmic reticulum. In their shape and size resemble lysosomes. Since spherosomes contain acid phosphatase, they are probably related to lysosomes. Some authors believe that spherosomes and lysosomes are equivalent to each other, but most likely only in origin and shape. There is an assumption about their participation in the synthesis of fats (A. Frey-Wissling).
Ribosomes - very small organoids, their diameter is about 250A. In shape they are almost spherical. Some of them are attached to the outer membranes of the endoplasmic reticulum, some of them are in a free state in the cytoplasm. A cell can contain up to 5 million ribosomes. Ribosomes are found in chloroplasts and mitochondria, where they synthesize part of the proteins of which these organoids are built, and the enzymes that function in them.
The main function is the synthesis of specific proteins according to information coming from the nucleus. Their composition: protein and ribosomal ribonucleic acid (RNA) in equal proportions. Their structure is small and large subunits formed from ribonucleotide.
Microtubules.Microtubules - peculiar derivatives of the endoplasmic reticulum. Found in many cells. Their name itself speaks of their shape - one or two, located in parallel, tubules with a cavity inside. Outside diameter within 250A. The walls of microtubules are built from protein molecules. Spindle filaments are formed from microtubules during cell division.
Core
The nucleus was discovered in a plant cell by R. Brown in 1831. It is located in the center of the cell or near the cell membrane, but is surrounded by cytoplasm on all sides. In most cases, there is one nucleus in the cell, several nuclei are in the cells of some algae, as well as fungi. Non-cellular green algae has hundreds of nuclei. Multinucleated cells of non-articulated lacteras. There are no nuclei in the cells of bacteria and blue-green algae.
The shape of the core is most often close to the shape of a ball or ellipse. Depends on the shape, age and function of the cell. In the meristematic cell, the nucleus is large, round in shape and occupies 3/4 of the cell volume. In parenchymal cells of the epidermis having a large central vacuole, the nucleus has a lenticular shape and is pushed along with the cytoplasm to the periphery of the cell. This is a sign of a specialized, but already aging cell. A cell lacking a nucleus can only live for a short time. The nuclear-free cells of the sieve tubes are living cells, but they do not live long. In all other cases, nuclear-free cells are dead.
The core has a double shell, through the pores in which the contents
nuclei (nucleoplasm) can communicate with the contents of the cytoplasm. Membrane membranes of the nucleus are equipped with ribosomes and communicate with the membranes of the endoplasmic reticulum of the cell. One or two nucleoli and chromosomes are located in the nucleoplasm. Nucleoplasm is a colloidal system of Zola, reminiscent of a thickened gelatin in consistency. According to Russian biochemists (I. Zbarsky, et al.), The core contains four fractions of proteins: simple proteins - globulins 20%, deoxyribonucleoproteins - 70%, acid proteins - 6% and residual proteins 4%. They are localized in the following nuclear structures: DNA proteides (alkaline proteins) in the chromosomes, RNA proteids (acidic proteins) in the nucleoli, partly in the chromosomes (during the synthesis of messenger RNA) and in the nuclear membrane. Globulins form the basis of nucleoplasm. Residual proteins (nature not specified) form a nuclear membrane.
The bulk of the core proteins are complex alkaline proteins of deoxyribonucleoproteins, which are based on DNA.
DNA molecule.DNA molecule - polynucleotide and consists of nucleotides. The nucleotide contains three components: a sugar molecule (deoxyribose), a nitrogenous molecule, and a phosphoric acid molecule. Deoxyribose is connected with the nitrogenous base glycosidic, and with phosphoric acid - an ether bond. In DNA, in various combinations there are only 4 varieties of nucleotides that differ from each other in nitrogenous bases. Two of them (adenine and guanine) belong to purine nitrogen compounds, and cytosine and thymine to pyrimidine. DNA molecules are not located on the same plane, but consist of two spiral strands, i.e. two parallel chains twisted one around the other form one DNA molecule. They are bonded to each other via a hydrogen bond between nitrogenous bases, and the purine bases of one chain attach the pyrimidine bases of the other (Fig. 14). The structure and chemistry of the DNA molecule was discovered by English (Crick) and American (Watson) scientists and published in 1953. This point is considered to be the beginning of the development of molecular genetics. The molecular weight of DNA is 4-8 million. The number of nucleotides (various options) is up to 100 thousand. The DNA molecule is very stable, its stability is ensured by the fact that throughout its length it has the same thickness - 20A (8A - the width of the pyrimidine base + 12A - the width of the purine base). If radioactive phosphorus is introduced into the body, then the label will be detected in all phosphorus-containing compounds, except DNA (Levy, Sickewitz).
DNA molecules are carriers of heredity, because their structure encodes information on the synthesis of specific proteins that determine the properties of the body. Changes can occur under the influence of mutagenic factors (radioactive radiation, potent, chemical agents, alkaloids, alcohols, etc.).
RNA molecule.Ribonucleic acid molecules (RNA) significantly fewer DNA molecules. These are single chains of nucleotides. There are three types of RNA: ribosomal, the longest, forming numerous loops, information (matrix) and transport, the shortest. Ribosomal RNA is localized in the ribosomes of the endoplasmic reticulum and accounts for 85% of the total RNA of the cell.
Informational RNA in its structure resembles a leaf of clover. Its amount is 5% of the total RNA in the cell. It is synthesized in the nucleoli. Its assembly occurs in chromosomes during the interphase period. Its main function is the transfer of information from DNA to ribosomes, where protein synthesis takes place.
Transport RNA, as it is now established, is a whole family of compounds related in structure and biological function. Each living cell, according to a rough estimate, contains 40-50 individual transport RNAs and their total number in nature, given the species differences, is huge. (Acad. V. Engelhardt). They are called transport because their molecules are engaged in transport maintenance of the intracellular process of protein synthesis. Combining with free amino acids, they deliver them to the ribosomes in the protein chain under construction. These are the smallest RNA molecules, composed on average of 80 nucleotides. Localized in the matrix of cytoplasm and make up about 10% of cellular RNA
RNA contains four nitrogenous bases, but in contrast to DNA, uracil is in place of thymine instead of thymine.
The structure of chromosomes.Chromosomes were first discovered in the late 19th century by the classics of cytology Fleming and Strasburger (1882, 1884), and the Russian cell researcher I.D. Chistyakov discovered them in 1874.
The main structural element of chromosomes is the nucleus. They have a different shape. These are either straight or curved sticks, oval bodies, balls, the sizes of which vary.
Depending on the location of the centromere, direct, equal, and unequal chromosomes are distinguished. The internal structure of chromosomes is shown in Fig. 15, 16. It should be noted that the deoxyribonucleoprotein is a monomer of the chromosome.
In the chromosome of deoxyribonucleoproteins, 90-92%, of which 45% is DNA and 55% is protein (histone). In a small amount in the chromosome, RNA is also represented (informational).
The chromosome also has a pronounced transverse structure - the presence of thickened areas - discs, which were back in 1909. were called genes. This term was proposed by the Danish scientist Johansen. In 1911, the American scientist Morgan proved that genes are the main hereditary units and they are distributed in the chromosomes in a linear order and, therefore, the chromosome has qualitatively different sections. In 1934, the American scientist Pinter proved the discontinuity of the morphological structure of chromosomes and the presence of disks in chromosomes, and disks are places where DNA accumulates. This served as the beginning of the creation of chromosomal maps, which indicated the location (locus) of the location of the gene that determines one or another trait of the organism. A gene is a DNA double helix site that contains information about the structure of a single protein. This is a section of a DNA molecule that determines the synthesis of a single protein molecule. DNA does not directly participate in protein synthesis. It only contains and stores information about the structure of the protein.
The DNA structure, consisting of several thousand consecutively located 4 nucleotides, is a code of heredity.
The code of heredity. Protein synthesis.The first message on the DNA code was made by the American biochemist Nirenberg in 1961 in Moscow at the International Biochemical Congress. The essence of the DNA code is as follows. Each amino acid corresponds to a portion of the DNA chain of three adjacent nucleotides (triplets). So, for example, a site consisting of TT-T (a triplet of 3 thymine-containing nucleotides) corresponds to the amino acid lysine, a triplet A (adenine) - C (cytosine) - A (adenine) - cysteine, etc. Suppose that a gene is represented by a chain of nucleotides arranged in the following order: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G. Having broken this series into triplets, we will immediately decipher which amino acids and in what order will be located in the synthesized protein.
The number of possible combinations of 4 available nucleotides of three is 4 × 64. Based on these ratios, the number of different triplets in excess will be enough to provide information on the synthesis of numerous proteins that determine both the structure and functions of the body. For protein synthesis, an exact copy of this information is sent to the ribosomes in the form of messenger RNA. In addition to i-RNA, a large number of molecules of various transport ribonucleic acids (t-RNAs), ribosomes, and a number of enzymes are involved in decoding and synthesis. Each of the 20 amino acids binds to T-RNA - a molecule with a molecule. Each of the 20 amino acids has its own t-RNA. T-RNAs have chemical groups capable of "recognizing" their amino acid, choosing it from the available amino acids. This happens with the help of special enzymes. Having recognized its amino acid, t-RNA enters into a compound with it. A ribosome is attached to the beginning of the chain (molecule) of i-RNA, which, advancing along the i-RNA, connects with each other exactly those amino acids whose order is encoded by the nucleotide sequence of this I-RNA. As a result, a protein molecule is formed, the composition of which is encoded in one of the genes.
Nucleoli- an integral structural part of the core. These are spherical bodies. They are very changeable, change their shape and structure, appear and disappear. There are one, two. For each plant a certain number. The nuclei disappear when the cell prepares for division, and then reappear; they are apparently involved in the synthesis of ribonucleic acids. If the nucleolus is destroyed by a focused beam of x-ray or ultraviolet rays, then cell division is suppressed.
The role of the nucleus in cell life. The nucleus serves as the controlling center of the cell - it directs cellular activity and contains heredity carriers (genes) that determine the characteristics of a given organism. The role of the nucleus can be identified if, using microsurgical techniques, remove it from the cell and observe the consequence of this. A series of experiments proving an important role in the regulation of cell growth was carried out by Hemmerling on the unicellular green alga Acetobularia. This seaweed reaches a height of 5 cm, looks like a mushroom, has the likeness of “roots” and “legs”. Above ends with a large disk-shaped "hat". The cell of this alga has one nucleus located in the basal part of the cell.
Hammerling found that if the leg is cut, the lower part continues to live and the cap is completely regenerated after the operation. The upper part, devoid of the nucleus, survives for some time, but, in the end, dies, not being able to restore the lower part. Consequently, the nucleus of acetobularia is necessary for the metabolic reactions underlying growth.
The nucleus promotes cell wall formation. This can be illustrated by experiments with the alga Voucheria and Spyrogyra. By releasing the contents of cells from cut filaments into water, we can get lumps of cytoplasm with one, with several nuclei and without nuclei. In the first two cases, the cell membrane formed normally. In the absence of a core, the shell did not form.
In the experiments of I.I. Gerasimov (1890) with spirogyra, it was found that cells with a double nucleus double the length and thickness of the chloroplast. In nuclear-free cells, the process of photosynthesis continues, assimilation starch is formed, but the process of its hydrolysis is attenuated, which is explained by the absence of hydrolytic enzymes that can be synthesized in ribosomes only according to the DNA of the nucleus. The life of a protoplast without a core is inferior and short-lived. In the experiments of I.I. Gerasimov’s nuclear-free spirogyra cells lived 42 days and died. One of the most important functions of the nucleus is to supply the cytoplasm with ribonucleic acid, which is necessary for the synthesis of protein in the cell. Removal of the nucleus from the cell leads to a gradual decrease in the content of RNA in the cytoplasm and a slowdown in protein synthesis in it.
The most important role of the nucleus is in the transfer of traits from cell to cell, from organism to organism, and it does this in the process of nuclear fission and the cell as a whole.
Cell division.Cells multiply by division. In this case, two daughter cells with the same set of hereditary material enclosed in chromosomes as the mother cell are formed from one cell. In somatic cells, chromosomes are represented by two, the so-called homologous chromosomes, in which allelic genes (carriers of opposite traits, for example, white and red pea petals, etc.), traits of two parental pairs are embedded. In this regard, in the somatic cells of the plant body, there is always a double set of chromoses, denoted 2p. Chromosomes have a pronounced personality. The quantity and quality of chromosomes is a characteristic feature of each species. So, in the strawberry cells the diploid set of chromoses is 14, (2n), the apple tree is 34, the Jerusalem artichoke is 102, etc.
Mitosis (karyokinesis) - somatic cell division was first described by E. Russov (1872) and I.D. Chistyakov (1874). Its essence lies in the fact that two daughter cells with the same set of chromoses are formed from the mother cell by division. The cell cycle is composed of interphase and mitosis proper. The method of microautoradiography found that the longest and most difficult is the interphase - the period of the "resting" core, because during this period there is a doubling of nuclear material. Interphase is divided into three phases:
Q1 - presynthetic (its duration is 4-6 hours);
S - synthetic (10-20 hours);
Q2 - postsynthetic (2-5 hours).
During the Q1 phase, preparations are underway for DNA reduplication. And in the S phase, DNA is reduced, the cell doubles the supply of DNA. In the Q2 phase, the enzymes and structures necessary to trigger mitosis are formed. Thus, in the interphase, DNA molecules in the chromosomes are split into two identical strands, and information RNA is assembled on their matrix. The latter carries information about the structure of specific proteins in the cytoplasm, and in the nucleus, each of the DNA strands completes the missing half of its molecule. In this process of doubling (reduction), a unique DNA feature is manifested, which consists in the ability of DNA to accurately reproduce itself. The resulting daughter DNA molecules are automatically obtained by exact copies of the parent molecule, because during reduplication, complementary (АТ; Г-Ц; etc.) bases from the environment join each half.
In the prophase of mitotic division, doubled chromosomes become noticeable. In metaphase, they are all located in the equatorial zone, located in one row. Spindle threads are formed (from microtubules connecting to each other). The shell of the nucleus and nucleolus disappear. Thickened chromosomes are split lengthwise into two daughter chromosomes. This is the essence of mitosis. It provides an accurate distribution of doubled DNA molecules between daughter cells. This ensures the transmission of hereditary information encrypted in DNA.
In anaphase, daughter chromosomes begin to move to opposite poles. In the center, the first fragments of the cell membrane (fragmoblast) appear.
In the telophase, nuclei are formed in daughter cells. The contents of the mother cell (organelle) is distributed between the resulting daughter cells. The cell membrane is completely formed. This ends the cytokinesis (Fig. 17).
Meiosis - Reduction Division was discovered and described in the 90s of the last century by V.I. Belyaev. The essence of the division is that from a somatic cell containing a 2n (double, diploid) chromosome set, four haploid cells are formed, with "n", a half set of chromoses. This type of division is complex and consists of two stages. The first is chromosome reduction. Doubled chromosomes are located in the equatorial zone in pairs (two parallel homologous chromosomes). At this moment, conjugation (adhesion) by chromosis, crossing-over (cross) can occur, and as a result, an exchange of chromosis sites occurs. As a result of this, part of the genes of paternal chromosomes becomes part of maternal chromoses and vice versa. The appearance of both chromosomes as a result of this does not change, but their qualitative composition becomes different. Paternal and maternal heredities are redistributed and mixed.
In the anaphase of meiosis, homologous chromosomes with the help of spindle filaments diverge at the poles, at which, after a short rest period (the filaments disappear, but the septum between the new nuclei does not form), the mitosis process begins - a metaphase in which all chromosomes are located in one plane and their longitudinal splitting occurs on daughter chromosomes. With anaphase of mitosis with the help of a spindle, they diverge at the poles, where four nuclei form, and as a result, four haploid cells. In the cells of some tissues during their development under the influence of certain factors, incomplete mitosis occurs and the number of chromoses in the nuclei doubles due to the fact that they do not diverge at the poles. As a result of such violations of a natural or artificial nature, tetraploids and polyploids arise. With the help of meiosis, germ cells are formed - gametes, as well as spores, elements of sexual and asexual reproduction of plants (Fig. 18).
Amitosis is a direct fission of the nucleus. With amitosis, a spindle of division is not formed and the shell of the nucleus does not decompose, as in mitosis. Previously, amitosis was considered as a primitive form of division. It has now been established that it is associated with the degradation of the body. It is a simplified version of a more complex fission of the kernel. Amitosis occurs in the cells and tissues of nucellus, endosperm, tuber parenchyma, leaf petioles, etc.
Mitochondria - It is a spiral, round, elongated or branched organelle.
The concept of mitochondria was first proposed by Benda in 1897. Mitochondria can be detected in living cells using phase contrast and interference microscopy in the form of grains, granules or filaments. These are quite mobile structures that can move, merge with each other, share. When stained with special methods in dead cells under light microscopy, the mitochondria look like small grains (granules), diffusely distributed in the cytoplasm or concentrated in some specific zones.
As a result of the destruction of glucose and fats in the presence of oxygen, energy is generated in the mitochondria, and organic matter is converted into water and carbon dioxide. In this way, animal organisms receive the basic energy necessary for life. Energy is accumulated in adenosine triphosphate (ATP), and more precisely, in its macroergic bonds. The function of mitochondria is closely related to the oxidation of organic compounds and the use of energy released during their decay for the synthesis of ATP molecules. Therefore, mitochondria are often called cell energy stations, or organelles of cellular respiration. ATP acts as an energy supplier, transferring one of its energy-rich terminal phosphate groups to another molecule, and turns into ADP.
It is believed that in evolution, mitochondria were prokaryotic microorganisms that became symbiotics in the body of an ancient cell. Subsequently, they became vitally necessary, which was associated with an increase in the oxygen content in the Earth’s atmosphere. On the one hand, mitochondria removed excess toxic oxygen for the cell, and on the other hand, they provided energy.
Without mitochondria, the cell is practically unable to use oxygen as a substance that provides energy supply, and can only meet its energy needs through anaerobic processes. Thus, oxygen is poison, but poison is vital for the cell, and excess oxygen is just as harmful as its deficiency.
Mitochondria can change their shape and move to those areas of the cell where the need for them is highest. So, in cardiomyocytes, mitochondria are located near myofibrils, in the cells of the renal tubules near basal protrusions, etc. The cell contains up to a thousand mitochondria, and their number depends on the activity of the cell.
Mitochondria have an average transverse size of 0.5 ... 3 microns. Depending on the size, small, medium, large and giant mitochondria are distinguished (they form a branched network - the mitochondrial reticulum). The size and number of mitochondria are closely related to the activity of the cell and its energy consumption. They are extremely variable and, depending on cell activity, oxygen content, hormonal influences, can swell, change the number and structure of cristae, vary in number, shape and size, as well as enzymatic activity.
The bulk density of mitochondria, the degree of development of their inner surface, and other indicators depend on the energy needs of the cell. In lymphocytes there are only a few mitochondria, and in the liver cells there are 2 ... 3 thousand.
Mitochondria consist of a matrix, an inner membrane, a perimitochondrial space, and an outer membrane. The outer mitochondrial membrane separates the organelle from the hyaloplasm. Usually it has smooth contours and is closed so that it is a membrane bag.
The outer membrane from the inner is separated by a perimitochondrial space with a width of about 10 ... 20 nm. The inner mitochondrial membrane limits the actual inner content of mitochondria - the matrix. The inner membrane forms numerous protrusions inward of the mitochondria, which have the appearance of flat ridges, or cristae.
The cristae can take the form of plates (trabecular) and tubules (multivesicular at the cut) in shape, and they are directed longitudinally or transversely with respect to mitochondria.
Each mitochondria is filled with a matrix, which in electron micrographs looks denser than the surrounding cytoplasm. The mitochondrial matrix is \u200b\u200bhomogeneous (homogeneous), sometimes fine-grained, of various electron densities. Thin filaments with a thickness of about 2 ... 3 nm and granules with a size of about 15 ... 20 nm are revealed in it. Matrix filaments are DNA molecules, and small granules are mitochondrial ribosomes. The matrix contains enzymes, one single-stranded, cyclic DNA, mitochondrial ribosomes, many Ca 2+ ions.
The autonomous system of protein synthesis of mitochondria is represented by histone-free DNA molecules. DNA is short, has the shape of a ring (cyclic) and contains 37 genes. Unlike nuclear DNA, there are practically no non-coding nucleotide sequences in it. Features of the structure and organization bring together mitochondrial DNA with bacterial cell DNA. On the mitochondrial DNA, various types of RNA molecules are synthesized: informational, transfer (transport) and ribosomal. Information RNA of mitochondria is not subjected to splicing (cutting out sections that do not carry information load). The small size of the mitochondrial DNA molecules cannot determine the synthesis of all mitochondrial proteins. Most mitochondrial proteins are under the genetic control of the cell nucleus and are synthesized in the cytoplasm, since mitochondrial DNA is weakly expressed and can provide the formation of only part of the oxidative phosphorylation chain enzymes. Mitochondrial DNA encodes no more than ten proteins that are localized in membranes and are structural proteins responsible for the correct integration of individual functional protein complexes of mitochondrial membranes. Proteins that carry out transport functions are also synthesized. Such a system of protein synthesis does not provide all the functions of mitochondria, therefore autonomy of mitochondria is limited and relative.
In mammals, mitochondria during fertilization are transmitted only through the egg, and sperm introduces nucleus DNA into the new organism.
In the matrix of mitochondria, ribosomes are formed that differ from cytoplasmic ribosomes. They participate in the synthesis of a number of mitochondrial proteins not encoded by the nucleus. Mitochondrial ribosomes have a sedimentation number of 60 (in contrast to cytoplasmic with a sedimentation number of 80). The sedimentation number is the deposition rate during centrifugation and ultracentrifugation. By structure, mitochondrial ribosomes are close to the ribosomes of prokaryotic organisms, but are smaller and are sensitive to certain antibiotics (chloramphenicol, tetracycline, etc.).
The inner mitochondrial membrane has a high degree of selectivity in the transport of substances. Enzymes of the oxidative phosphorylation chain, electron carrier proteins, ATP, ADP, pyruvate, and other transport systems closely adjacent to each other are attached to its inner surface. As a result of the close arrangement of enzymes on the inner membrane, high conjugation (interconnection) of biochemical processes is ensured, which increases the rate and the efficiency of catalytic processes.
Electron microscopy reveals mushroom particles protruding into the lumen of the matrix. They have ATP-synthetic (forms ATP from ADP) activity. Electron transport takes place along the respiratory chain localized in the inner membrane, which contains four large enzyme complexes (cytochromes). When electrons pass through the respiratory chain, hydrogen ions are pumped from the matrix into the perimitochondrial space, which ensures the formation of a proton gradient (pump). The energy of this gradient (differences in the concentration of substances and the formation of membrane potential) is used for the synthesis of ATP and the transport of metabolites and inorganic ions. The carrier proteins contained in the inner membrane transport organic phosphates, ATP, ADP, amino acids, fatty acids, tri- and dicarboxylic acids through it.
The outer mitochondrial membrane is more permeable to low molecular weight substances, since there are many hydrophilic protein channels in it. Specific receptor complexes are located on the outer membrane, through which proteins from the matrix are transported into the perimitochondrial space.
In terms of its chemical composition and properties, the outer membrane is close to other intracellular membranes and plasmolemma. It contains enzymes that metabolize fats, activate (catalyze) the conversion of amines, amino oxidase. If the enzymes of the outer membrane remain active, then this is an indicator of the functional safety of mitochondria.
There are two autonomous subcompartments in the mitochondria. The perimitochondrial space, or the outer mitochondrial chamber (external subcompartment), is formed due to the penetration of protein complexes of the hyaloplasm, while the internal subcompartment (mitochondrial matrix) is partially formed due to the synthetic activity of mitochondrial DNA. The internal sub-compartment (matrix) contains DNA, RNA, and ribosomes. It has a high level of Ca 2+ ions in comparison with hyaloplasm. Hydrogen ions accumulate in the external sub-compartment. The enzymatic activity of the external and internal subcompartments, the composition of the proteins are very different. The internal sub-compartment has a higher electron density than the external.
Specific mitochondrial markers are cytochrome oxidase and succinate dehydrogenase enzymes, the detection of which allows the quantitative characterization of energy processes in mitochondria.
The main function of mitochondria - ATP synthesis. Initially, in the hyaloplasm, sugars (glucose) are destroyed to lactic and pyruvic acids (pyruvate) with the simultaneous synthesis of a small amount of ATP. As a result of glycolysis of one glucose molecule, two ATP molecules are used, and four are formed. Thus, only two ATP molecules make up the positive balance. These processes occur without oxygen (anaerobic glycolysis).
All subsequent stages of energy production occur in the process of aerobic oxidation, which provides the synthesis of a large amount of ATP. In this case, organic matter is destroyed to CO 2 and water. Oxidation is accompanied by the transfer of protons to their acceptors. These reactions are carried out using a number of enzymes of the tricarboxylic acid cycle, which are located in the mitochondrial matrix.
Electron transfer systems and its associated phosphorylation of ADP (oxidative phosphorylation) are built into the cristal membranes. In this case, the transfer of electrons from one electron acceptor protein to another takes place and, finally, their binding to oxygen, as a result of which water is formed. At the same time, part of the energy released during such oxidation in the electron transfer chain is stored as a macroergic bond during ADP phosphorylation, which leads to the formation of a large number of ATP molecules, the main intracellular energy equivalent. The process of oxidative phosphorylation occurs on the membranes of mitochondrial cristae using the oxidation chain proteins and the ADP ATP synthetase phosphorylation enzyme located here. As a result of oxidative phosphorylation, 36 ATP molecules are formed from one glucose molecule.
To some hormones and substances on the mitochondria there are specialized (affinity) receptors. Triiodothyronine normally accelerates the synthetic activity of mitochondria. Interleukin-1 and high concentrations of triiodothyronine disconnect the oxidative phosphorylation chains, cause mitochondrial swelling, which is accompanied by an increase in the formation of thermal energy.
New mitochondria are formed by division, constriction or budding. In the latter case, protomitochondria are formed, gradually increasing in size.
Protomitochondria is a small organelle with outer and inner membranes. The inner membrane does not have or contains underdeveloped cristae. Organella is characterized by low levels of aerobic phosphorylation. With the formation of a constriction, the contents of the mitochondria are distributed between two new fairly large organelles. With any method of reproduction, each of the newly formed mitochondria has its own genome.
Older mitochondria are destroyed by autolysis (self-digestion by the cell using lysosomes) to form autolysosomes. From the autolysosome, a residual body is formed. With complete digestion, the contents of the residual body, consisting of low molecular weight organic substances, are excreted by exocytosis. With incomplete digestion, the remains of mitochondria can accumulate in the cell in the form of layered bodies or granules with nipofuscin. Insoluble calcium salts accumulate in the part of mitochondria with the formation of crystals - calcifications. The accumulation of mitochondrial degeneration products can lead to cell dystrophy.
The structure and function of mitochondria is a rather complex issue. The presence of organelles is characteristic of almost all nuclear organisms - both for autotrophs (plants capable of photosynthesis) and for heterotrophs, which are almost all animals, some plants and fungi.
The main purpose of mitochondria is the oxidation of organic substances and the subsequent use of the energy released as a result of this process. For this reason, organelles also have a second (unofficial) name - cell energy stations. They are sometimes called "catabolism plastids."
What is mitochondria
The term is of Greek origin. Translated, this word means "thread" (mitos), "seed" (chondrion). Mitochondria are permanent organoids that are of great importance for the normal functioning of cells and make it possible for the whole organism to exist.

"Stations" have a specific internal structure, which varies depending on the functional state of the mitochondria. Their shape can be of two types - oval or oblong. The latter often has a branching appearance. The number of organelles in one cell ranges from 150 to 1500.
A special case is germ cells. Only one spiral organelle is present in the sperm, while female gametes contain hundreds of thousands more mitochondria. In the cell, organelles are not fixed in one place, but can move along the cytoplasm, combine with each other. Their size is 0.5 microns, the length can reach 60 microns, while the minimum figure is 7 microns.

Determining the size of one “power station” is not an easy task. The fact is that when viewed in an electron microscope, only part of the organelle gets into the slice. It so happens that spiral mitochondria has several sections that can be taken for separate, independent structures.
Only a three-dimensional image will make it possible to find out the exact cellular structure and to understand whether we are talking about 2-5 separate organoids or about one with a complex form of mitochondria.
Structural features
The mitochondrial membrane consists of two layers: external and internal. The latter includes various outgrowths and folds that have a leaf-like and tubular shape.
Each membrane has a special chemical composition, a certain amount of certain enzymes and a specific purpose. The outer shell from the inner is separated by an intermembrane space with a thickness of 10-20 nm.
The structure of the organelle in the figure with the captions looks very clear.

The structure of the mitochondria
Looking at the structure diagram, the following description can be made. The viscous space inside the mitochondria is called the matrix. Its composition creates a favorable environment for the flow of necessary chemical processes in it. It contains microscopic granules that promote reactions and biochemical processes (for example, accumulate glycogen ions and other substances).
The matrix contains DNA, coenzymes, ribosomes, t-RNA, inorganic ions. ATP synthase and cytochromes are located on the surface of the inner layer of the shell. Enzymes contribute to processes such as the Krebs cycle (CTK), oxidative phosphorylation, etc.
Thus, the main task of the organoid is performed by both the matrix and the inner side of the shell.
Mitochondrial function
The purpose of the “power plants” can be characterized by two main tasks:
- energy production: oxidation processes are carried out in them with the subsequent release of ATP molecules;
- storage of genetic information;
- participation in the synthesis of hormones, amino acids and other structures.
The process of oxidation and energy production takes place in several stages:


Schematic drawing of ATP synthesis
It is worth noting: as a result of the Krebs cycle (citric acid cycle), ATP molecules are not formed, molecules are oxidized and carbon dioxide is released. This is an intermediate step between glycolysis and the electron transport chain.
Table "Functions and structure of mitochondria"

What determines the number of mitochondria in a cell
The prevailing number of organoids accumulates near those parts of the cell where there is a need for energy resources. In particular, a large number of organelles are collected in the area of \u200b\u200bmyofibrils, which are part of the muscle cells that ensure their contraction.

In male germ cells, structures are localized around the flagellum axis - it is assumed that the need for ATP is due to the constant movement of the tail of the gamete. The arrangement of mitochondria in protozoa, which use special cilia for movement, looks like organelles accumulate under the membrane at their base in exactly the same way.
As for nerve cells, the localization of mitochondria is observed near the synapses through which signals of the nervous system are transmitted. In cells synthesizing proteins, organelles accumulate in the ergastoplasma zones - they supply energy that provides this process.
Who discovered mitochondria
The cell structure gained its name in 1897-1898 thanks to C. Brand. Otto Wagburg was able to prove the relationship of cellular respiration with mitochondria in 1920.
Conclusion
Mitochondria are the most important component of a living cell, acting as an energy station that produces ATP molecules, thereby ensuring the processes of cell activity.
The work of mitochondria is based on the oxidation of organic compounds, resulting in the generation of energy potential.
Outer membrane
Inner membrane
Matrix m-on, matrix, krista. it has smooth contours, does not form protrusions or folds. It accounts for about 7% of the area of \u200b\u200ball cell membranes. Its thickness is about 7 nm, it is not connected with any other membranes of the cytoplasm and is closed to itself, so it is a membrane bag. The outer membrane is separated from the inner intermembrane space about 10-20 nm wide. The inner membrane (about 7 nm thick) limits the actual inner content of mitochondria,
its matrix or mitoplasm. A characteristic feature of the inner membrane of mitochondria is their ability to form numerous protrusions inside the mitochondria. Such protrusions most often have the form of flat ridges, or cristae. The distance between the membranes in the crista is about 10-20 nm. Often cristae can branch or form finger-shaped processes, bend and not have a pronounced orientation. In protozoa, unicellular algae, in some cells of higher plants and animals, outgrowths of the inner membrane look like tubes (tubular cristae).
The mitochondrial matrix has a fine-grained homogeneous structure, sometimes thin filaments collected in a ball (about 2-3 nm) and granules about 15-20 nm are detected in it. Now it has become known that mitochondrial matrix strands are DNA molecules in the composition of the mitochondrial nucleoid, and small granules are mitochondrial ribosomes.
Mitochondrial function
1. In mitochondria ATP synthesis takes place (see. Oxidative phosphorylation)
Intermembrane pH ~ 4, matrix pH ~ 8 | the protein content in m: 67% is the matrix, 21% is the discovery of the mine, 6% is the inside of the mine and 6% is in the inter-mine
Distemper - a single mitochondrial system
outer m-na: porin-pores allow passage up to 5 kD | inner m-na: cardiolipin-impervious m-well for ions |
international production: enzyme groups phosphorylate nucleotides and nucleotide sugars
Inland:
matrix: metabolic enzymes - lipid oxidation, carbohydrate oxidation, tricarboxylic acid cycle, Krebs cycle
Origin from bacteria: the amoeba Pelomyxa palustris does not contain unity from eukaryotes, lives in symbiosis with aerobic bacteria | own DNA | OX-like processes
Mitochondrial DNA
Myochondria division
replicated
in interphase | replication is not associated with S-phase | during the cl cycle, the mitochis are once divided in two, forming a constriction, the constriction first on the inside m-not | ~ 16.5 kb | ring encodes 2 rRNA 22 tRNA and 13 proteins |
protein transport: signal peptide | amphiphilic curl | mitochondrial recognition receptor |
Oxidative Phosphorylation
Electron transfer circuit
ATP synthase
in liver cells, m live ~ 20 days mitochondrial division by the formation of constriction
16569 bp \u003d 13 proteins, 22tRNA, 2pRNA | smooth outer m-na (porins - protein permeability up to 10 kDa) folded inner (crista) m-na (75% proteins): transport carrier proteins, f-you, components of the respiratory chain and ATP synthase, cardiolipin) matrix ( enriched with f-tami citrate cycle) inter 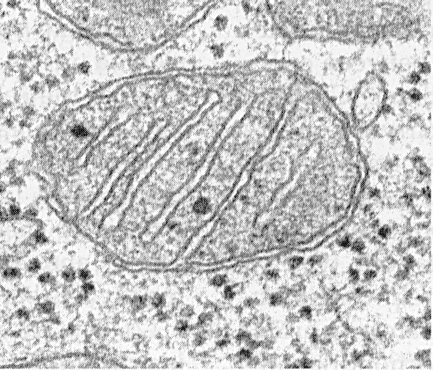
Mitochondria or chondriosomes (from the Greek. Mitos - thread, chondrion - grain, soma - body) are granular or filamentous organelles present in the cytoplasm of protozoa, plants and animals. Mitochondria can be observed in living cells, since they have a fairly high density. In living cells, mitochondria can move, move, merge with each other.
In different species, the sizes of mitochondria are very variable, as well as their shape is variable (Fig. 199). Nevertheless, in most cells, the thickness of these structures is relatively constant (about 0.5 μm), and the length fluctuates, reaching up to 7-60 μm in filamentous forms.
Studying the size and number of mitochondria is not such a simple matter. This is due to the fact that the size and number of mitochondria that are visible on ultra-thin sections do not correspond to reality.
Conventional calculations show that about 200 mitochondria are present in the liver cell. This represents more than 20% of the total cytoplasm and about 30-35% of the total amount of protein in the cell. The surface area of \u200b\u200ball mitochondria of the liver cell is 4-5 times larger than the surface of its plasma membrane. Most mitochondria are found in oocytes (about 300,000) and in the giant amoeba Chaos chaos (up to 500,000).
In green plant cells, the number of mitochondria is less than in animal cells, since part of their functions can be performed by chloroplasts.
The localization of mitochondria in the cells is different. Typically, mitochondria accumulate near those areas of the cytoplasm where there is a need for ATP formed in mitochondria. So, in skeletal muscle, mitochondria are located near myofibrils. In spermatozoa, mitochondria form a spiral case around the flagellum axis; this is probably due to the need to use ATP for sperm tail movement. Similarly, in protozoa and in other cells equipped with cilia, mitochondria are localized directly under the cell membrane at the base of the cilia, which require ATP to work. In the axons of nerve cells, mitochondria are located near the synapses, where the process of transmission of a nerve impulse occurs. In secretory cells that synthesize large amounts of proteins, mitochondria are closely associated with ergastoplasma zones; they probably supply ATP for the activation of amino acids and protein synthesis on ribosomes.
Ultrastructure of mitochondria.
Mitochondria, regardless of their size or shape, have a universal structure, their ultrastructure is uniform. Mitochondria are limited to two membranes (Fig. 205). The outer mitochondrial membrane separates it from the hyaloplasm, it has smooth contours, does not form protrusions or folds, and the thickness is about 7 nm. It accounts for about 7% of the area of \u200b\u200ball cell membranes. The membrane is not connected with any other cytoplasmic membranes, is closed on itself and is a membrane bag. The outer membrane from the inner is separated by an intermembrane space with a width of about 10-20 nm. The inner membrane (about 7 nm thick) limits the actual internal content of the mitochondria, its matrix or mitoplasm. The inner mitochondrial membrane forms numerous intrusions inside the mitochondria. Such protrusions most often have the form of flat ridges, or cristae.
The total surface of the inner mitochondrial membrane in the liver cell is approximately one third of the surface of all cell membranes. The mitochondria of the heart muscle cells contain three times as many cristae as the liver mitochondria, which reflects the differences in the functional loads of the mitochondria of different cells. The distance between the membranes in the crista is about 10-20 nm.
Mitochondrial cristae extending from the inner membrane and extending toward the matrix do not completely block the mitochondrial cavity and do not violate the continuity of the matrix filling it.
The orientation of the cristae with respect to the long axis of the mitochondria is different for different cells. Orientation can be perpendicular (liver, kidney cells) cristae; in the heart muscle, a longitudinal arrangement of cristae is observed. Crista can branch or form finger-shaped processes, bend and not have a pronounced orientation (Fig. 208). In protozoa, unicellular algae, in some cells of higher plants and animals, outgrowths of the inner membrane look like tubes (tubular cristae).
The mitochondrial matrix has a fine-grained homogeneous structure in which DNA molecules are detected in the form of thin filaments collected in a ball (about 2-3 nm) and mitochondrial ribosomes in the form of granules about 15-20 nm in size. The deposits of magnesium and calcium salts in the matrix form large (20-40 nm) dense granules.
The functions of mitochondria.
Mitochondria carry out the synthesis of ATP, which occurs as a result of oxidation of organic substrates and phosphorylation of ADP.
The initial stages of carbohydrate oxidation are called anaerobic oxidation, or glycolysis and occur in the hyaloplasm and do not require oxygen. The oxidation substrate during anaerobic energy production are hexoses, and primarily glucose; some bacteria have the ability to extract energy by oxidizing pentoses, fatty acids or amino acids.
In glucose, the amount of potential energy contained in the bonds between the C, H and O atoms is about 680 kcal per 1 mol (i.e., 180 g glucose).
In a living cell, this huge amount of energy is released in the form of a stepwise process, controlled by a number of oxidizing enzymes, and is not associated with the transition of chemical bond energy into heat, as during combustion, but passes into a macroenergetic bond in ATP molecules, which are synthesized using released energy from ADP and phosphate.
Trioses formed as a result of glycolysis, primarily pyruvic acid, are involved in mitochondria in further oxidation. In this case, the energy of cleavage of all chemical bonds is used, which leads to the release of CO 2, to the consumption of oxygen and the synthesis of a large amount of ATP. These processes are associated with the oxidation cycle of tricarboxylic acids and with the respiratory chain of electron transfer, where ADP phosphorylation and the synthesis of cellular “fuel” and ATP molecules occur (Fig. 209).
In the tricarboxylic acid cycle (the Krebs cycle, or the citric acid cycle), pyruvate formed as a result of glycolysis first loses the CO 2 molecule and, being oxidized to acetate (a two-carbon compound), combines with coenzyme A. Then, acetyl coenzyme A combines with oxalacetate (a four-carbon compound), forms six-carbon citrate (citric acid). Then there is a cycle of oxidation of this six-carbon compound to four-carbon oxalacetate, again binding to acetyl coenzyme A, and then the cycle repeats. During this oxidation, two CO 2 molecules are released, and the electrons released during the oxidation are transferred to the acceptor coenzyme molecules (NAD-nicotinamide adenine dinucleotide), which further involve them in the electron transfer chain. Therefore, in the cycle of tricarboxylic acids there is no ATP synthesis itself, but there is an oxidation of molecules, electron transfer to acceptors and emission of CO 2. All the events described above within the mitochondria occur in their matrix.
Oxidation of the initial substrate leads to the release of CO 2 and water, but thermal energy is not released, as during combustion, but ATP molecules are formed. They are synthesized by another group of proteins that are not directly related to oxidation. In the inner mitochondrial membranes, large protein complexes, enzymes, ATP synthetases are located on the surface of the membranes that look into the matrix. In an electron microscope, they are visible in the form of the so-called “mushroom” bodies, which completely line the surface of the membranes, looking into the matrix. The bodies have a leg and a head, with a diameter of 8-9 nm. Consequently, both the oxidizing chain enzymes and ATP synthesis enzymes are localized in the inner mitochondrial membranes (Fig. 201b).
The respiratory chain is the main system for converting energy in mitochondria. Here, sequential oxidation and restoration of the elements of the respiratory chain occurs, as a result of which energy is released in small portions. Due to this energy, ATP is formed from ADP and phosphate at three points of the chain. Therefore, they say that oxidation (electron transfer) is associated with phosphorylation (ADP + Fn → ATP, i.e., oxidative phosphorylation occurs.
The energy released during electron transport is stored in the form of a proton gradient on the membrane. It turned out that during electron transfer in the mitochondrial membrane, each complex of the respiratory chain directs oxidation free energy to move protons (positive charges) through the membrane, from the matrix to the intermembrane space, which leads to the formation of potential differences on the membrane: positive charges prevail in the intermembrane space, and negative - from the side of the mitochondrial matrix. When the potential difference (220 mV) is reached, the protein complex of ATP synthetase begins to transport protons back to the matrix, while it turns one form of energy into another: it forms ATP from ADP and inorganic phosphate. This is the combination of oxidative processes with synthetic processes, with phosphorylation of ADP. While substrates are oxidizing, while protons are pumped through the inner mitochondrial membrane, ATP synthesis associated with this occurs, i.e. oxidative phosphorylation occurs.
These two processes can be separated. In this case, electron transfer continues, as well as substrate oxidation, but ATP synthesis does not occur. In this case, the energy released during oxidation is converted into thermal energy.
Oxidative phosphorylation in bacteria.
In prokaryotic cells capable of oxidative phosphorylation, the elements of the tricarboxylic acid cycle are localized directly in the cytoplasm, and the respiratory chain and phosphorylation enzymes are associated with the cell membrane, with its protrusions protruding inside the cytoplasm, with the so-called mesosomes (Fig. 212). It should be noted that such bacterial mesosomes can be associated not only with the processes of aerobic respiration, but also in some species participate in cell division, in the process of DNA distribution in new cells, in the formation of the cell wall, etc.
On the plasma membrane in the mesosomes of some bacteria, conjugated processes of both oxidation and ATP synthesis are carried out. Spherical particles similar to those found in the mitochondria of eukaryotic cells were found in an electron microscope in the fractions of the plasma membranes of bacteria. Thus, in bacterial cells capable of oxidative phosphorylation, the plasma membrane plays a role similar to the inner mitochondrial membrane of eukaryotic cells.
An increase in the number of mitochondria.
Mitochondria can increase their numbers especially with cell division or with an increase in the functional load of the cell. There is a constant renewal of mitochondria. For example, in the liver, the average lifespan of mitochondria is about 10 days.
An increase in the number of mitochondria occurs through the growth and division of previous mitochondria. This assumption was first made by Altman (1893), who described mitochondria under the term “bioblasts”. It is possible to observe in vivo division, fragmentation of long mitochondria into shorter ones by constriction, which resembles the binary method of division of bacteria.
A real increase in the number of mitochondria by division was established when studying the behavior of mitochondria in living cells of tissue culture. During the cell cycle, mitochondria grow to several microns, and then fragment, divide into smaller bodies.
Mitochondria can merge with each other and multiply according to the principle: mitochondria from mitochondria.
Autoproduction of mitochondria.
Two-membrane organelles have a complete system of auto-reproduction. In mitochondria and plastids, there is DNA on which information, transfer and ribosomal RNAs and ribosomes are synthesized, which carry out the synthesis of mitochondrial and plastid proteins. However, these systems, although autonomous, are limited in their capabilities.
DNA in mitochondria is a cyclic molecule without histones and thus resembles bacterial chromosomes. Their size is about 7 microns, 16-19 thousand nucleotide pairs of DNA enter into one cyclic molecule of animal mitochondria. In humans, mitochondrial DNA contains 16.5 thousand n.p., it is completely deciphered. It was found that the mitochondral DNA of various objects is very homogeneous, their difference lies only in the magnitude of introns and non-transcribed sites. All mitochondrial DNAs are multiple copies assembled into groups, clusters. So in one mitochondria of the rat liver can contain from 1 to 50 cyclic DNA molecules. The total amount of mitochondrial DNA per cell is about one percent. The synthesis of mitochondrial DNA is not related to the synthesis of DNA in the nucleus.
As in bacteria, mitochondral DNA is collected in a separate zone - the nucleoid, its size is about 0.4 microns in diameter. In long mitochondria, there can be from 1 to 10 nucleoids. When dividing a long mitochondria, a section containing a nucleoid is separated from it (similar to the binary division of bacteria). The amount of DNA in individual mitochondrial nucleoids can fluctuate 10 times depending on the type of cells.
In some cultures, in cells from 6 to 60% of mitochondria do not have a nucleoid, which can be explained by the fact that the division of these organelles is more likely associated with fragmentation, and not with the distribution of nucleoids.
As already mentioned, mitochondria can both divide and merge with each other. When mitochondria merge with each other, an exchange of their internal components can occur.
It is important to emphasize that rRNA and ribosomes of mitochondria and cytoplasm are sharply different. If 80s ribosomes are found in the cytoplasm, then the mitochondrial ribosomes of plant cells belong to 70s ribosomes (consist of 30s and 50s subunits, contain 16s and 23s RNAs characteristic of prokaryotic cells), and smaller ribosomes are found in animal cell mitochondria (about 50s).
Mitochondrial ribosomal RNAs are synthesized on mitochondrial DNA. In the mitoplasm on the ribosomes is the synthesis of proteins. It stops, unlike synthesis on cytoplasmic ribosomes, under the action of the antibiotic chloramphenicol, which inhibits the synthesis of protein in bacteria.
On the mitochondrial genome, 22 transport RNAs are synthesized. The triplet code of the mitochondrial synthetic system is different from that used in hyaloplasm. Despite the presence of seemingly all the components necessary for protein synthesis, small mitochondrial DNA molecules cannot encode all mitochondrial proteins, only a small fraction of them. So DNA of 15 thousand n.p. can encode proteins with a total molecular weight of about 6x10 5. At the same time, the total molecular weight of the proteins of the particles of the complete respiratory ensemble of mitochondria reaches a value of about 2x10 6.
Given that in addition to oxidative phosphorylation proteins, mitochondria also include tricarboxylic acid cycle enzymes, DNA and RNA synthesis enzymes, amino acid activation enzymes, and other proteins, it can be seen that in order to encode these numerous proteins and rRNA and tRNA, the amount of genetic information in the short mitochondrial DNA molecule is clearly not enough. Decoding the nucleotide sequence of human mitochondrial DNA showed that it encodes only 2 ribosomal RNAs, 22 transfer RNAs, and a total of 13 different polypeptide chains.
It has now been proven that most of the mitochondrial proteins are under genetic control from the cell nucleus and are synthesized outside the mitochondria. Most mitochondrial proteins are synthesized on ribosomes in the cytosol. These proteins have special signal sequences that are recognized by receptors on the outer mitochondrial membrane. These proteins can be inserted into them (see the analogy with peroxisome membrane), and then move to the inner membrane. This transfer occurs at the contact points of the outer and inner membranes where such transport is noted. Most mitochondrial lipids are also synthesized in the cytoplasm.
All this indicates the endosymbiotic origin of mitochondria, that mitochondria are organisms such as bacteria that are in symbiosis with a eukaryotic cell.
Chondriom.
The totality of all mitochondria in one cell is called a chondrioma. It can be different depending on the type of cells. In many cells, chondriomas consist of scattered numerous mitochondria uniformly located throughout the cytoplasm or localized in groups at the sites of intense ATP expenditure. In both of these cases, mitochondria function alone, their cooperative work, possibly coordinated by some signals from the cytoplasm. There is a completely different type of chondrioma, when instead of small single scattered mitochondria, one giant branched mitochondria is located in the cell.
Such mitochondria are found in unicellular green algae (for example, Chlorella). They form a complex mitochondrial network or mitochondrial reticulum (Reticulum miyochondriale). According to the chemo osmotic theory, the biological meaning of the appearance of such a giant branched mitochondrial structure, united by its external and internal membranes, is that ATP synthesis can occur at any point on the surface of the inner membrane of such a branched mitochondria, which will go anywhere in the cytoplasm, where There is a need for this.
In the case of giant branched mitochondria, at any point on the inner membrane, a potential sufficient to start the synthesis of ATP can accumulate. From these positions, the mitochondrial reticulum is, as it were, an electrical conductor, a cable connecting the distant points of such a system. The mitochondrial reticulum proved to be very useful not only for small mobile cells, such as chlorella, but also for larger structural units such as, for example, myofibrils in skeletal muscle.
It is known that skeletal muscle consists of a mass of muscle fibers, symplasts containing many nuclei. The length of such muscle fibers reaches 40 microns, with a thickness of 0.1 microns - this is a gigantic structure containing a great many myofibrils, all of which contract simultaneously, synchronously. For contraction, a large amount of ATP, which mitochondria provide at the level of z-disks, is delivered to each contraction unit, to myofibril. On the longitudinal ultrathin sections of skeletal muscles, an electron microscope shows numerous rounded small sections of mitochondria located in the vicinity of sarcomeres. Muscular mitochondria are not small balls or rods, but rather arachnid structures, the processes of which branch and extend over long distances, sometimes across the entire diameter of the muscle fiber.
At the same time, mitochondrial branching surrounds each myofibril in the muscle fiber, supplying them with the ATP necessary for muscle contraction. Therefore, in the plane of the z-disk, mitochondria represent a typical mitochondrial reticulum. Such a layer or floor of the mitochondrial reticulum is repeated twice for each sarcomere, and the entire muscle fiber has thousands of transverse “floor” layers of the mitochondrial reticulum. Between the "floors" along the myofibrils are filamentous mitochondria connecting these mitochondrial layers. Thus, a three-dimensional picture of the mitochondrial reticulum, passing through the entire volume of muscle fiber, was created.
It was further established that between the branches of the mitochondrial reticulum and the filiform longitudinal mitochondria there are special intermitochondrial compounds or contacts (MMC). They are formed by tightly adjacent outer mitochondrial membranes of contacting mitochondria, the intermembrane space and membranes in this zone have an increased electron density. Through these special formations, a functional union of neighboring mitochondria and mitochondrial reticulums takes place into a single, cooperative energy system. All myofibrils in the muscle fiber are reduced synchronously along their entire length, therefore, the supply of ATP on any part of this complex machine should also occur synchronously, and this can only happen if a huge number of branched mitochondria-conductors are connected with each other using contacts.
The fact that intermitochodial contacts (MMK) are involved in the energy association of mitochondria with each other was possible on cardiomyocytes, heart muscle cells.
Chondriomas of heart muscle cells do not form branching structures, but are represented by many small elongated mitochondria located without special order between myofibrils. However, all neighboring mitochondria join each other using mitochondrial contacts of the same type as in skeletal muscle, only their number is very large: on average, there are 2-3 MMCs per mitochondria that bind mitochondria into a single chain, where each link has chain (Streptio mitochondriale) is a separate mitochondria.
It turned out that intermitochondrial contacts (MMC), as a mandatory structure of heart cells, were found in cardiomyocytes of both the ventricles and atria of all vertebrates: mammals, birds, reptiles, amphibians and bony fish. Moreover, MMCs were found (but in a smaller number) in the heart cells of some insects and mollusks.
The amount of MMC in cardiomyocytes varies depending on the functional load on the heart. The number of MMC increases with increasing physical activity of animals and, conversely, when the load on the heart muscle decreases, there is a sharp decrease in the number of MMC.







