ในฐานะที่เป็นโครงสร้างภายในของไมโตคอนเดรียจะเรียกว่า โครงสร้างและหน้าที่ของไมโตคอนเดรียและพลาสดิด
mitochondria
ในเซลล์ของเนื้อเยื่อสัตว์พบไมโตคอนเดรียในปี 1882 และในพืชเท่านั้นในปี 1904 (ในอับเรณูของลิลลี่น้ำ) มันเป็นไปได้ที่จะสร้างฟังก์ชั่นทางชีวภาพหลังจากการแยกและการทำให้บริสุทธิ์ของเศษส่วนโดยการปั่นแยกเศษส่วน พวกเขามีโปรตีน 70% และไขมันประมาณ 30%, RNA และ DNA, วิตามิน A, B 6, B 12, K, E, กรดโฟลิกและแพนโทธีนิก, ไรโบฟลาวิน, เอนไซม์ต่างๆ Mitochondria มีเมมเบรนสองชั้นด้านนอกจะแยกออร์แกเนลออกจากไซโตพลาสซึมและรูปแบบด้านในมีจำนวนมากกว่าคริสต้า ช่องว่างทั้งหมดระหว่างเยื่อหุ้มเซลล์จะเต็มไปด้วยเมทริกซ์ (รูปที่ 13)
หน้าที่หลักของไมโตคอนเดรียคือการมีส่วนร่วมในการหายใจของเซลล์ บทบาทของไมโตคอนเดรียในการหายใจถูกจัดตั้งขึ้นในปี 2493-2494 ระบบเอนไซม์ที่ซับซ้อนของวงจร Krebs นั้นมีความเข้มข้นในเยื่อหุ้มชั้นนอก ในระหว่างการออกซิเดชั่นของพื้นผิวการหายใจพลังงานจะถูกปล่อยออกมาซึ่งจะสะสมในโมเลกุล ADP ที่เกิดขึ้นทันทีส่วนใหญ่คือ ATP ในระหว่างการออกซิเดทีฟฟอสโฟรีเลชันที่เกิดขึ้นใน cristae พลังงานที่เก็บไว้ในสารประกอบมาโครกิกจะถูกใช้ในอนาคตเพื่อตอบสนองความต้องการทั้งหมดของเซลล์
การก่อตัวของไมโทคอนเดรียในเซลล์เกิดขึ้นอย่างต่อเนื่องจากจุลินทรีย์มักเกิดขึ้นเกี่ยวข้องกับความแตกต่างของโครงสร้างเยื่อหุ้มเซลล์ พวกเขาสามารถคืนค่าในเซลล์โดยการหารและพวกเขาออกมา Mitochondria ไม่คงทนอายุขัยของพวกเขาคือ 5-10 วัน
Mitochondria เป็นสถานีพลังงาน“ เซลล์” พวกเขารวมพลังงานซึ่งถูกเก็บไว้ใน "สะสม" ของพลังงาน - โมเลกุล ATP และไม่กระจายตัวในเซลล์ การละเมิดโครงสร้างของไมโทคอนเดรียนำไปสู่การหยุดชะงักของกระบวนการทางเดินหายใจและเป็นผลให้พยาธิวิทยาของร่างกาย
เครื่อง Golgiเครื่อง Golgi (คำพ้องความหมาย - dictiosomes) เป็นสแต็คของ 3-12 แบนปิดล้อมล้อมรอบด้วยแผ่นเมมเบรนสองแผ่นที่เรียกว่าถังจากขอบของฟองอากาศจำนวนมากที่ปลดออก (300-500) ความกว้างของถังคือ 6-90 A ความหนาของเยื่อหุ้มคือ 60-70 A
อุปกรณ์ Golgi เป็นศูนย์กลางในการสังเคราะห์การสะสมและการแยกโพลีแซคคาไรด์โดยเฉพาะเซลลูโลสและมีส่วนร่วมในการกระจายและการขนส่งโปรตีนภายในเซลล์รวมถึงการก่อตัวของแวคิวโอโลโซม ในเซลล์พืชเป็นไปได้ที่จะติดตามการมีส่วนร่วมของอุปกรณ์ Golgi ในลักษณะของแผ่นมัธยฐานและการเจริญเติบโตของเยื่อหุ้มเซลล์เพคโต้เซลลูโลส
อุปกรณ์ Golgi ได้รับการพัฒนามากที่สุดในช่วงระยะเวลาของเซลล์ที่ใช้งานอยู่ เมื่อมันมีอายุมากขึ้นก็ค่อยๆเสื่อมถอยไปและหายไป
lysosomeslysosomes - มีขนาดค่อนข้างเล็ก (เส้นผ่าศูนย์กลางประมาณ 0.5 ไมครอน) พวกเขาถูกเคลือบด้วยเมมเบรนโปรตีน lipoid เนื้อหาของไลโซโซมเป็นเอนไซม์ไฮโดรไลติกจำนวนมากที่ทำหน้าที่ของการย่อยภายในเซลล์ (lysis) ของโปรตีน macromolecules, กรดนิวคลีอิก, โพลีแซคคาไรด์ หน้าที่หลักของพวกเขาคือการย่อยของแต่ละส่วนของเซลล์โปรโตพลาสต์ (autophagy - self-devouring) กระบวนการนี้ดำเนินต่อเนื่องจาก phagocytosis หรือ pinocytosis บทบาททางชีวภาพของกระบวนการนี้เป็นสองเท่า ประการแรกมันป้องกันเนื่องจากการขาดแคลนชั่วคราวของเซลล์อะไหล่เซลล์ช่วยชีวิตเนื่องจากโปรตีนตามรัฐธรรมนูญและสารอื่น ๆ และประการที่สองมีการปล่อยจากอวัยวะส่วนเกินหรือสวมใส่ (plastids, ไมโทคอนเดรีย ฯลฯ ) เยื่อ lysosome ป้องกันการปล่อยเอนไซม์เข้าสู่ไซโตพลาสซึม มิฉะนั้นมันจะถูกย่อยโดยเอนไซม์เหล่านี้
ในเซลล์ที่ตายแล้วไลโซโซมจะถูกทำลายเอนไซม์อยู่ในเซลล์และเนื้อหาทั้งหมดจะถูกย่อย สิ่งที่เหลืออยู่คือเปลือกเพคโตเซลลูโลส
Lysosomes เป็นผลิตภัณฑ์ของกิจกรรมของอุปกรณ์ Golgi, ถุงที่แยกออกจากมัน, ซึ่งเอนไซม์ย่อยสลาย organoid สะสมนี้
Sferosomy - ร่างกาย - โปรตีน lipoid กลม 0.3-0.4 ไมครอน ในทุกโอกาสมันเป็นอนุพันธ์ของอุปกรณ์ Golgi หรือเอนโดพลาสซึมย้อนกลับ ในรูปร่างและขนาดคล้ายไลโซโซม เนื่องจาก spherosomes มี acid phosphatase จึงอาจเกี่ยวข้องกับ lysosomes ผู้เขียนบางคนเชื่อว่า spherosomes และ lysosomes เทียบเท่ากัน แต่มีแนวโน้มมากที่สุดในแหล่งกำเนิดและรูปร่าง มีข้อสันนิษฐานเกี่ยวกับการมีส่วนร่วมในการสังเคราะห์ไขมัน (A. Frey-Wissling)
ไรโบโซม - ออร์การอยด์ขนาดเล็กมากเส้นผ่านศูนย์กลางอยู่ที่ 250A พวกมันเกือบจะเป็นทรงกลม บางส่วนถูกยึดติดกับเยื่อหุ้มชั้นนอกของเอนโดพลาสซึมเรติเคิลซึ่งบางส่วนอยู่ในสถานะอิสระในไซโตพลาสซึม เซลล์สามารถมีไรโบโซมสูงถึง 5 ล้าน ไรโบโซมพบได้ในคลอโรพลาสต์และไมโทคอนเดรียซึ่งพวกเขาสังเคราะห์ส่วนของโปรตีนที่ทำขึ้นเป็นสารอินทรีย์และเอนไซม์ที่ทำงานในพวกเขา
หน้าที่หลักคือการสังเคราะห์โปรตีนเฉพาะตามข้อมูลจากนิวเคลียส องค์ประกอบของพวกเขา: โปรตีนและกรด ribosomal ribonucleic (RNA) ในสัดส่วนที่เท่ากัน โครงสร้างของพวกมันคือหน่วยย่อยขนาดเล็กและขนาดใหญ่ที่เกิดขึ้นจากไรโบนิวคลีโอไทด์
microtubulesmicrotubules - อนุพันธ์ที่แปลกประหลาดของเอนโดพลาสซึมเรติเคิล พบได้หลายเซลล์ ชื่อของพวกเขาเองพูดถึงรูปร่างของพวกเขา - หนึ่งหรือสองตั้งอยู่ในแบบคู่ขนาน tubules กับโพรงภายใน เส้นผ่านศูนย์กลางภายนอก 250A ผนังของ microtubules สร้างขึ้นจากโมเลกุลโปรตีน ไส้แกนหมุนเกิดขึ้นจาก microtubules ระหว่างการแบ่งเซลล์
แกนกลาง
นิวเคลียสถูกค้นพบในเซลล์พืชโดย R. Brown ในปี 1831 มันตั้งอยู่ในใจกลางของเซลล์หรือใกล้กับเยื่อหุ้มเซลล์ แต่ถูกล้อมรอบด้วยไซโตพลาสซึมในทุกด้าน ในกรณีส่วนใหญ่มีเซลล์หนึ่งนิวเคลียสในนิวเคลียสหลายเซลล์อยู่ในเซลล์ของสาหร่ายบางชนิดรวมถึงเชื้อรา สาหร่ายสีเขียวที่ไม่ใช่เซลลูล่าร์มีนิวเคลียสนับร้อย เซลล์หลายเซลล์ของแลคเทอร่าที่ไม่ได้ประกบ ไม่มีนิวเคลียสในเซลล์ของแบคทีเรียและสาหร่ายสีน้ำเงินแกมเขียว
รูปร่างของแกนมักจะใกล้เคียงกับรูปร่างของลูกหรือวงรี ขึ้นอยู่กับรูปร่างอายุและหน้าที่ของเซลล์ ในเซลล์ meristematic นิวเคลียสมีขนาดใหญ่กลมเป็นรูปร่างและมีปริมาตรของเซลล์ 3/4 ในเซลล์เนื้อเยื่อของหนังกำพร้าที่มีแวคิวโอลกลางขนาดใหญ่นิวเคลียสมีรูปร่างเป็นแม่และถูกย้ายไปพร้อมกับไซโทพลาสซึมไปยังรอบนอกของเซลล์ นี่คือสัญญาณของเซลล์ที่มีความเชี่ยวชาญ เซลล์ที่ขาดนิวเคลียสจะมีชีวิตอยู่ได้เพียงช่วงเวลาสั้น ๆ เท่านั้น เซลล์ที่ปราศจากนิวเคลียร์ของหลอดตะแกรงเป็นเซลล์ที่มีชีวิต แต่จะมีชีวิตอยู่ไม่นาน ในกรณีอื่นทั้งหมดเซลล์ปลอดนิวเคลียร์ตาย
แกนมีเปลือกสองชั้นผ่านรูขุมขนที่มีเนื้อหา
นิวเคลียส (nucleoplasm) สามารถสื่อสารกับเนื้อหาของไซโตพลาสซึม เมมเบรนของนิวเคลียสมีการติดตั้งไรโบโซมและสื่อสารกับเยื่อหุ้มเซลล์ของเอนโดพลาสซึมเรติเคิลของเซลล์ ในนิวคลีโอพลาสซึมจะมีนิวคลีโอลีและโครโมโซมหนึ่งหรือสองตัว นิวคลีโอพลาสซึมเป็นระบบคอลลอยด์ของ Zola ซึ่งคล้ายกับเจลาตินที่มีความหนา ตามที่นักชีวเคมีชาวรัสเซีย (I. Zbarsky และอื่น ๆ ) แกนประกอบด้วยสี่ส่วนของโปรตีน: โปรตีนง่าย - โกลบูลิ 20%, deoxyribonucleoproteins - 70%, โปรตีนกรด - 6% และโปรตีนที่เหลือ 4% พวกมันถูกแปลในโครงสร้างนิวเคลียร์ต่อไปนี้: DNA proteides (โปรตีนอัลคาไลน์) ในโครโมโซม, โปรตีน RNA (โปรตีนกรด) ในนิวเคลียสส่วนหนึ่งในโครโมโซม (ในระหว่างการสังเคราะห์ Messenger RNA) และในเยื่อหุ้มนิวเคลียร์ โกลบูลินเป็นพื้นฐานของนิวเคลียส โปรตีนตกค้าง (ไม่ได้ระบุธรรมชาติ) ก่อตัวเป็นเยื่อหุ้มนิวเคลียร์
ส่วนใหญ่ของโปรตีนหลักคือโปรตีนอัลคาไลน์ที่ซับซ้อน deoxyribonucleoproteins ซึ่งอยู่บนพื้นฐานของดีเอ็นเอ
โมเลกุลดีเอ็นเอโมเลกุลดีเอ็นเอ - polynucleotide และประกอบด้วยนิวคลีโอไทด์ นิวคลีโอไทด์มีองค์ประกอบสามส่วน: โมเลกุลน้ำตาล (deoxyribose), โมเลกุลไนโตรเจนและโมเลกุลกรดฟอสฟอริก Deoxyribose เชื่อมต่อกับ glycosidic ฐานไนโตรเจนและกรดฟอสฟอริก - พันธะอีเธอร์ ใน DNA ในชุดต่าง ๆ มีนิวคลีโอไทด์เพียง 4 พันธุ์เท่านั้นที่แตกต่างจากนิวคลีโอไทด์ในฐานไนโตรเจน สองของพวกเขา (adenine และ guanine) เป็นของสารประกอบ purine ไนโตรเจนและ cytosine และ thymine เพื่อ pyrimidine โมเลกุลของ DNA ไม่ได้อยู่ในระนาบเดียวกัน แต่ประกอบด้วยเกลียวสองเส้นนั่นคือ โซ่คู่ขนานสองบิดหนึ่งรอบอีกรูปหนึ่งโมเลกุลดีเอ็นเอ พวกมันถูกผูกมัดซึ่งกันและกันโดยพันธะไฮโดรเจนระหว่างฐานไนโตรเจนกับฐาน purine ของห่วงโซ่หนึ่งเข้าร่วมฐาน pyrimidine ของอื่น ๆ (รูปที่ 14) โครงสร้างและเคมีของโมเลกุล DNA ถูกค้นพบโดยนักวิทยาศาสตร์ชาวอังกฤษ (Crick) และอเมริกัน (วัตสัน) และเผยแพร่ต่อสาธารณะในปี 1953 ประเด็นนี้ถือเป็นจุดเริ่มต้นของการพัฒนาพันธุศาสตร์โมเลกุล น้ำหนักโมเลกุลของ DNA อยู่ที่ 4-8 ล้านจำนวนนิวคลีโอไทด์ (ตัวเลือกต่าง ๆ ) นั้นสูงถึง 100,000 โมเลกุล DNA นั้นมีความเสถียรมากความเสถียรของมันนั้นมีความหนาเหมือนกันคือ 20A (8A - ความกว้างของฐาน pyrimidine + 12A - ความกว้างของฐาน purine) ถ้ามีการนำกัมมันตภาพรังสีฟอสฟอรัสเข้าสู่ร่างกายฉลากจะถูกตรวจพบในสารประกอบที่มีฟอสฟอรัสทั้งหมดยกเว้น DNA (Levy, Sickewitz)
โมเลกุลดีเอ็นเอเป็นพาหะของการถ่ายทอดทางพันธุกรรมเพราะ โครงสร้างของพวกเขาเข้ารหัสข้อมูลเกี่ยวกับการสังเคราะห์โปรตีนเฉพาะที่กำหนดคุณสมบัติของร่างกาย การเปลี่ยนแปลงสามารถเกิดขึ้นได้ภายใต้อิทธิพลของปัจจัยการก่อกลายพันธุ์ (กัมมันตภาพรังสีกัมมันตภาพรังสีสารเคมีแอลคาลอยด์แอลกอฮอล์ ฯลฯ )
โมเลกุลอาร์เอ็นเอโมเลกุลกรด Ribonucleic (RNA) โมเลกุลดีเอ็นเอน้อยลงอย่างมาก นี่เป็นโซ่เดี่ยวของนิวคลีโอไทด์ มีสามประเภทของ RNA: ribosomal ที่ยาวที่สุดก่อรูปหลายลูปข้อมูล (เมทริกซ์) และการขนส่งสั้นที่สุด Ribosomal RNA มีการแปลในไรโบโซมของเอนโดพลาสซึมเรติเคิลและคิดเป็น 85% ของ RNA ทั้งหมดของเซลล์
ให้ข้อมูล RNA ในโครงสร้างของมันคล้ายใบไม้ของโคลเวอร์ จำนวนของมันคือ 5% ของ RNA ทั้งหมดในเซลล์ มันถูกสังเคราะห์ในนิวเคลียส การชุมนุมของมันเกิดขึ้นในโครโมโซมในช่วงระยะเวลา หน้าที่หลักคือการถ่ายโอนข้อมูลจาก DNA ไปยังไรโบโซมซึ่งการสังเคราะห์โปรตีนเกิดขึ้น
Transport RNA ซึ่งเป็นที่ยอมรับกันในขณะนี้เป็นสารประกอบทั้งครอบครัวที่เกี่ยวข้องกับโครงสร้างและหน้าที่ทางชีวภาพ แต่ละเซลล์ที่มีชีวิตตามการประมาณคร่าวๆประกอบด้วยการขนส่ง RNA 40-50 ตัวและจำนวนทั้งหมดในธรรมชาติตามความแตกต่างของสายพันธุ์นั้นมีขนาดใหญ่มาก (Acad. V. Engelhardt) พวกเขาเรียกว่าการขนส่งเพราะโมเลกุลของพวกเขามีส่วนร่วมในการบำรุงรักษาการขนส่งกระบวนการ intracellular ของการสังเคราะห์โปรตีน เมื่อรวมกับกรดอะมิโนอิสระพวกมันจะส่งไปยังไรโบโซมในห่วงโซ่โปรตีนที่กำลังก่อสร้าง เหล่านี้เป็นโมเลกุล RNA ที่เล็กที่สุดประกอบด้วยค่าเฉลี่ยของนิวคลีโอไทด์ 80 มีการแปลในเมทริกซ์ไซโตพลาสซึมและประกอบด้วย RNA ของเซลล์ประมาณ 10%
อาร์เอ็นเอประกอบด้วยเบสสี่เบส แต่ไม่เหมือน DNA ยูราซิลแทนที่ไทมีนในอาร์เอ็นเอ
โครงสร้างของโครโมโซมโครโมโซมถูกค้นพบครั้งแรกในช่วงปลายศตวรรษที่ 19 โดยคลาสสิกของเซลล์วิทยาเฟลมมิ่งและสตราสบูร์กเกอร์ (1882, 1884) และนักวิจัยเซลล์รัสเซีย Chistyakov ค้นพบพวกเขาในปี 1874
องค์ประกอบโครงสร้างหลักของโครโมโซมคือนิวเคลียส พวกเขามีรูปร่างที่แตกต่าง เหล่านี้เป็นแท่งตรงหรือโค้งรูปวงรีลูกขนาดที่แตกต่างกันไป
ขึ้นอยู่กับตำแหน่งของ centromere โครโมโซมที่เท่ากันโดยตรงและไม่เท่ากันนั้นแตกต่างกัน โครงสร้างภายในของโครโมโซมแสดงในรูปที่ 15, 16. ควรสังเกตว่า deoxyribonucleoprotein เป็นโมโนเมอร์ของโครโมโซม
ในโครโมโซมของ deoxyribonucleoproteins 90-92% ซึ่ง 45% เป็น DNA และ 55% เป็นโปรตีน (ฮิสโตน) RNA (ข้อมูล) ยังแสดงในจำนวนเล็กน้อยในโครโมโซม
โครโมโซมยังมีโครงสร้างตามขวางที่เด่นชัดคือมีพื้นที่หนา - แผ่นดิสก์ซึ่งเกิดขึ้นในปี 1909 ถูกเรียกว่ายีน คำนี้นำเสนอโดยนักวิทยาศาสตร์ชาวเดนมาร์ก Johansen ในปีพ. ศ. 2454 มอร์แกนนักวิทยาศาสตร์ชาวอเมริกันได้พิสูจน์ว่ายีนเป็นหน่วยพันธุกรรมหลักและมีการแพร่กระจายในโครโมโซมตามลำดับเชิงเส้นดังนั้นโครโมโซมจึงมีส่วนที่แตกต่างกันในเชิงคุณภาพ ในปีพ. ศ. 2477 นักวิทยาศาสตร์ชาวอเมริกันพินได้พิสูจน์ความไม่ต่อเนื่องของโครงสร้างทางสัณฐานวิทยาของโครโมโซมและการมีอยู่ของดิสก์ในโครโมโซมและดิสก์เป็นสถานที่ที่ดีเอ็นเอสะสม สิ่งนี้ทำหน้าที่เป็นจุดเริ่มต้นของการสร้างแผนที่โครโมโซมซึ่งระบุตำแหน่ง (สถานที) ของตำแหน่งของยีนที่กำหนดลักษณะของสิ่งมีชีวิตอย่างน้อยหนึ่งอย่าง ยีนคือบริเวณที่มีลักษณะเป็นเกลียวคู่ของ DNA ที่มีข้อมูลเกี่ยวกับโครงสร้างของโปรตีนเดี่ยว นี่เป็นส่วนหนึ่งของโมเลกุลดีเอ็นเอที่กำหนดการสังเคราะห์โมเลกุลโปรตีนเดี่ยว DNA ไม่ได้มีส่วนร่วมในการสังเคราะห์โปรตีนโดยตรง มันมีเพียงและเก็บข้อมูลเกี่ยวกับโครงสร้างของโปรตีน
โครงสร้างดีเอ็นเอซึ่งประกอบด้วยนิวคลีโอไทด์ 4 พันที่อยู่ติดกันหลายพันเป็นรหัสของพันธุกรรม
รหัสของพันธุกรรม การสังเคราะห์โปรตีนข้อความแรกในรหัส DNA ถูกสร้างขึ้นโดย Nirenberg นักชีวเคมีชาวอเมริกันในปี 2504 ที่กรุงมอสโกในการประชุม Biochemical International สาระสำคัญของรหัส DNA มีดังนี้ กรดอะมิโนแต่ละตัวนั้นตรงกับส่วนของ DNA chain ของนิวคลีโอไทด์สามตัว ตัวอย่างเช่นไซต์ที่ประกอบด้วย TT-T (นิวคลีโอไทด์ที่มี 3 ไทมีน) สอดคล้องกับกรดอะมิโนไลซีน, triplet A (adenine) - C (ไซโตซิน) - A (adenine) - cysteine \u200b\u200bเป็นต้น สมมติว่ายีนมีสายโซ่ของนิวคลีโอไทด์เรียงตามลำดับต่อไปนี้: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G เมื่อแยกซีรีย์นี้ออกเป็นสามส่วนเราจะถอดรหัสกรดอะมิโนทันทีและในลำดับใดที่จะอยู่ในโปรตีนสังเคราะห์
จำนวนชุดค่าผสมที่เป็นไปได้ของนิวคลีโอไทด์ที่มีอยู่ 4 ตัวในสามตัวคือ 4 × 64 จากอัตราส่วนเหล่านี้จำนวนของแฝดสามที่แตกต่างกันจะเพียงพอที่จะให้ข้อมูลเกี่ยวกับการสังเคราะห์โปรตีนจำนวนมากที่กำหนดทั้งโครงสร้างและหน้าที่ของร่างกาย สำหรับการสังเคราะห์โปรตีนสำเนาที่แน่นอนของข้อมูลนี้จะถูกส่งไปยังไรโบโซมในรูปแบบของ Messenger RNA นอกเหนือจาก i-RNA แล้วโมเลกุลของกรดขนส่ง ribonucleic (t-RNAs), ไรโบโซมและเอนไซม์จำนวนมากเข้าร่วมในการถอดรหัสและการสังเคราะห์ กรดอะมิโนทั้ง 20 ชนิดนี้จับกับ T-RNA ซึ่งเป็นโมเลกุลที่มีโมเลกุล กรดอะมิโน 20 ตัวแต่ละตัวมี t-RNA ของตัวเอง T-RNA มีกลุ่มทางเคมีที่สามารถ "จดจำ" กรดอะมิโนของพวกเขาได้โดยเลือกจากกรดอะมิโนที่มีอยู่ สิ่งนี้เกิดขึ้นได้ด้วยความช่วยเหลือของเอนไซม์พิเศษ เมื่อรู้ว่ากรดอะมิโน t-RNA จะเข้าสู่สารประกอบด้วย ไรโบโซมติดอยู่ที่จุดเริ่มต้นของสายโซ่ (โมเลกุล) ของ i-RNA ซึ่งใกล้เคียงกับ i-RNA เชื่อมต่อกับกรดอะมิโนเหล่านั้นซึ่งมีลำดับเข้ารหัสโดยลำดับนิวคลีโอไทด์ของ I-RNA นี้ เป็นผลให้โมเลกุลโปรตีนถูกสร้างขึ้นซึ่งเป็นองค์ประกอบของการเข้ารหัสในหนึ่งในยีน
nucleoli- ส่วนโครงสร้างที่สำคัญของแกนกลาง เหล่านี้เป็นวัตถุทรงกลม พวกมันเปลี่ยนแปลงได้มากเปลี่ยนรูปร่างและโครงสร้างปรากฏและหายไป มีหนึ่งสอง สำหรับแต่ละโรงงานมีจำนวนที่แน่นอน นิวเคลียสจะหายไปเมื่อเซลล์เตรียมพร้อมสำหรับการแบ่งแล้วปรากฏขึ้นอีกครั้ง เห็นได้ชัดว่ามีส่วนร่วมในการสังเคราะห์กรด ribonucleic หากนิวเคลียสถูกทำลายโดยลำแสงโฟกัสของรังสีเอกซ์หรือรังสีอัลตราไวโอเลตการแบ่งเซลล์จะถูกระงับ
บทบาทของนิวเคลียสในชีวิตเซลล์ นิวเคลียสทำหน้าที่เป็นศูนย์กลางในการควบคุมของเซลล์ - มันทำหน้าที่ควบคุมกิจกรรมของเซลล์และมีผู้ให้บริการพันธุกรรม (พันธุกรรม) ที่กำหนดลักษณะของสิ่งมีชีวิตที่ได้รับ บทบาทของนิวเคลียสสามารถระบุได้ว่าหากใช้เทคนิคการผ่าตัดเล็ก ๆ ให้เอามันออกจากเซลล์และสังเกตผลที่ตามมาของสิ่งนี้ ชุดของการทดลองพิสูจน์บทบาทสำคัญในการควบคุมการเจริญเติบโตของเซลล์ดำเนินการโดย Hemmerling ใน Acetobularia สาหร่ายสีเขียวที่มีเซลล์เดียว สาหร่ายนี้มีความสูง 5 ซม. มีลักษณะเหมือนเห็ดมีความคล้ายคลึงกันของ "ราก" และ "ขา" ด้านบนจบลงด้วย "หมวก" รูปดิสก์ขนาดใหญ่ เซลล์ของสาหร่ายนี้มีนิวเคลียสหนึ่งอยู่ในส่วนฐานของเซลล์
Hammerling พบว่าหากตัดขาส่วนล่างจะยังคงอยู่และหมวกจะถูกสร้างใหม่อย่างสมบูรณ์หลังการผ่าตัด ส่วนบนไร้นิวเคลียสอยู่รอดมาได้ระยะหนึ่ง แต่สุดท้ายแล้วก็ตายไม่สามารถกู้คืนส่วนล่างได้ ดังนั้นนิวเคลียสของ acetobularia เป็นสิ่งจำเป็นสำหรับปฏิกิริยาการเผาผลาญพื้นฐานการเจริญเติบโต
นิวเคลียสส่งเสริมการสร้างผนังเซลล์ นี้สามารถแสดงได้โดยการทดลองกับ alga Voucheria และ Spyrogyra โดยการปล่อยเนื้อหาของเซลล์จากการตัดเส้นใยลงไปในน้ำเราสามารถรับไซโตพลาสซึมก้อนเดียวด้วยนิวเคลียสหลายชนิดและไม่มีนิวเคลียส ในสองกรณีแรกเยื่อหุ้มเซลล์เกิดขึ้นตามปกติ ในกรณีที่ไม่มีคอร์เชลล์จะไม่ก่อตัว
ในการทดลองของ I.I Gerasimov (1890) กับ spirogyra พบว่าเซลล์ที่มีนิวเคลียสสองเท่าของความยาวและความหนาของคลอโรพลาสต์ ในเซลล์ที่ปลอดนิวเคลียร์กระบวนการสังเคราะห์ด้วยแสงยังคงดำเนินต่อไปเกิดการดูดกลืนของแป้ง แต่กระบวนการของการไฮโดรไลซิสถูกลดทอนซึ่งอธิบายได้โดยการขาดเอนไซม์ไฮโดรไลติกที่สามารถสังเคราะห์ได้ในไรโบโซมตาม DNA ของนิวเคลียสเท่านั้น ชีวิตของโปรโตพลาสต์ที่ไม่มีแกนกลางนั้นด้อยกว่าและมีอายุสั้น ในการทดลองของ I.I เซลล์ spirogyra ที่ปลอดนิวเคลียร์ของ Gerasimov มีอายุ 42 วันแล้วตาย หนึ่งในหน้าที่ที่สำคัญที่สุดของนิวเคลียสคือการจัดหาไซโตพลาสซึมด้วยกรดริบบอนนิวคลีอิกซึ่งจำเป็นสำหรับการสังเคราะห์โปรตีนในเซลล์ การกำจัดนิวเคลียสออกจากเซลล์นำไปสู่การลดลงอย่างค่อยเป็นค่อยไปในเนื้อหาของ RNA ในไซโตพลาสซึมและการชะลอตัวของการสังเคราะห์โปรตีนในนั้น
บทบาทที่สำคัญที่สุดของนิวเคลียสคือการถ่ายโอนลักษณะจากเซลล์หนึ่งไปสู่อีกเซลล์หนึ่งจากสิ่งมีชีวิตไปสู่สิ่งมีชีวิตและมันทำเช่นนี้ในกระบวนการของการแตกตัวเป็นนิวเคลียสและเซลล์โดยรวม
การแบ่งเซลล์เซลล์คูณด้วยการหาร ในกรณีนี้เซลล์ลูกสาวสองเซลล์จะถูกสร้างขึ้นจากเซลล์หนึ่งซึ่งมีวัสดุทางพันธุกรรมชุดเดียวกันอยู่ในโครโมโซมเหมือนกับเซลล์แม่ ในโซมาติกเซลล์โครโมโซมจะถูกแทนด้วยสองโครโมโซมที่เรียกว่า homologous ซึ่งมียีนอัลลีลิกวางอยู่ (พาหะอักขระที่ตรงกันข้ามเช่นกลีบดอกถั่วสีขาวและสีแดง ฯลฯ ) สัญญาณของผู้ปกครองสองคู่ ในเรื่องนี้ในเซลล์ร่างกายของร่างกายพืชมักจะมีโครเมี่ยมเป็นสองเท่าซึ่งแสดงด้วย 2p โครโมโซมมีบุคลิกเด่นชัด ปริมาณและคุณภาพของโครโมโซมเป็นคุณลักษณะเฉพาะของสปีชีส์แต่ละชนิด ดังนั้นในเซลล์สตรอเบอร์รี่ชุดของโครโมโซมคือ 14, (2n), ต้นแอปเปิ้ลคือ 34, อาติโช๊คเยรูซาเล็มคือ 102, เป็นต้น
Mitosis (karyokinesis) - การแบ่งเซลล์ร่างกายได้รับการอธิบายครั้งแรกโดย E. Russov (1872) และ I.D. Chistyakov (1874) สาระสำคัญของมันอยู่ในความจริงที่ว่าเซลล์ลูกสาวสองคนที่มีชุดของโครโมโซมเดียวกันเกิดขึ้นจากการแบ่งเซลล์แม่โดยวงจรเซลล์ประกอบด้วย interphase และ mitosis ที่เหมาะสม วิธีการของ microautoradiography เป็นที่ยอมรับว่ายาวที่สุดและยากที่สุดคือ interphase - ระยะเวลาของนิวเคลียส "พักผ่อน" เนื่องจาก ในช่วงเวลานี้มีวัสดุนิวเคลียร์เพิ่มขึ้นเป็นสองเท่า เฟสจะแบ่งออกเป็นสามขั้นตอน:
Q1 - presynthetic (ระยะเวลา 4-6 ชั่วโมง)
S - สังเคราะห์ (10-20 ชั่วโมง)
Q2 - โพสต์สังเคราะห์ (2-5 ชั่วโมง)
ในช่วงไตรมาสที่ 1 มีการเตรียมการสำหรับการทำซ้ำดีเอ็นเอ และในเฟส S DNA จะลดลงเซลล์จะเพิ่มปริมาณดีเอ็นเอเป็นสองเท่า ในช่วงไตรมาสที่ 2 จะมีการสร้างเอนไซม์และโครงสร้างที่จำเป็นในการกระตุ้นการสร้างเซลล์ ดังนั้นในระหว่างเฟสโมเลกุลของดีเอ็นเอในโครโมโซมจะถูกแบ่งออกเป็นสองเส้นที่เหมือนกันและข้อมูล RNA ถูกประกอบเข้ากับเมทริกซ์ หลังถือข้อมูลเกี่ยวกับโครงสร้างของโปรตีนที่เฉพาะเจาะจงในพลาสซึมและในนิวเคลียสแต่ละเส้นดีเอ็นเอเสร็จสมบูรณ์ครึ่งหนึ่งของโมเลกุลที่ขาดหายไป ในกระบวนการของการเสแสร้ง (การลดลง) นี้คุณลักษณะที่เป็นเอกลักษณ์ของ DNA นั้นแสดงออกมาซึ่งประกอบไปด้วยความสามารถของ DNA ในการสืบพันธุ์ตัวเองได้อย่างถูกต้อง โมเลกุลดีเอ็นเอของลูกสาวที่ได้รับนั้นจะได้รับโดยอัตโนมัติจากสำเนาของโมเลกุลพ่อแม่เพราะ ในระหว่างการทำซ้ำการทำซ้ำฐาน (АТ; Г-Ц; ฯลฯ ) ฐานจากสิ่งแวดล้อมเข้าร่วมแต่ละครึ่ง
ในการทำนายของการแบ่งทิคติกโครโมโซมสองเท่าจะสังเกตเห็นได้ชัดเจน ใน metaphase ทั้งหมดอยู่ในเขตเส้นศูนย์สูตรตั้งอยู่ในแถวเดียว ไส้แกนหมุนเกิดขึ้น (จาก microtubules เชื่อมต่อกัน) เปลือกของนิวเคลียสและนิวเคลียสหายไป โครโมโซมหนาถูกแบ่งตามยาวออกเป็นสองโครโมโซมลูกสาว นี่คือสาระสำคัญของ mitosis ให้การกระจายที่แม่นยำของโมเลกุลดีเอ็นเอสองเท่าระหว่างเซลล์ลูกสาว สิ่งนี้ทำให้มั่นใจได้ว่าการถ่ายทอดข้อมูลทางพันธุกรรมที่เข้ารหัสใน DNA
ใน Anaphase โครโมโซมลูกสาวเริ่มย้ายไปที่ขั้วตรงข้าม ชิ้นส่วนแรกของเยื่อหุ้มเซลล์ปรากฏขึ้นตรงกลาง (ชิ้นส่วน)
ใน telophase นิวเคลียสจะเกิดขึ้นในเซลล์ลูกสาว เนื้อหาของเซลล์แม่ (organelle) มีการกระจายระหว่างเซลล์ลูกสาวที่เกิด เยื่อหุ้มเซลล์เกิดขึ้นอย่างเต็มที่ สิ่งนี้จะทำให้ cytokinesis เสร็จสมบูรณ์ (รูปที่ 17)
ไมโอซิส - กองลด ถูกค้นพบและอธิบายในยุค 90 ของศตวรรษที่แล้วโดย V.I. Belyaev สาระสำคัญของการแบ่งคือจากเซลล์โซมาติกที่ประกอบด้วยชุดโครเมียม 2n (double, diploid) จะมีการสร้างเซลล์ haploid สี่เซลล์โดยมี "n" ครึ่งชุด chromoses การแบ่งประเภทนี้มีความซับซ้อนและประกอบด้วยสองขั้นตอน อย่างแรกคือการลดโครโมโซม โครโมโซมสองเท่าตั้งอยู่ในเขตเส้นศูนย์สูตรเป็นคู่ (โครโมโซมคล้ายคลึงกันสองขนาน) ในขณะนี้การผันคำกริยา (การยึดเกาะ) โดย chromosis การข้ามข้าม (crossover) สามารถเกิดขึ้นได้และส่งผลให้เกิดการแลกเปลี่ยนไซต์ chromosis เป็นผลมาจากสิ่งนี้ส่วนหนึ่งของยีนของโครโมโซมพ่อกลายเป็นส่วนหนึ่งของโครโมโซมของมารดาและในทางกลับกัน การปรากฏตัวของโครโมโซมทั้งสองเป็นผลมาจากสิ่งนี้ไม่เปลี่ยนแปลง แต่องค์ประกอบเชิงคุณภาพของพวกเขาจะแตกต่างกัน การถ่ายทอดทางพันธุกรรมของพ่อและแม่จะถูกแจกจ่ายและผสม
ใน Anaphase ของไมโอซิส, โครโมโซมที่เหมือนกันด้วยความช่วยเหลือของแกนหมุนใยแก้วนำแสงแยกที่เสาซึ่งเกิดขึ้นหลังจากระยะเวลาพักสั้น ๆ (เส้นใยหายไป แต่เยื่อบุโพรงระหว่างนิวเคลียสใหม่ไม่ได้เกิดขึ้น) กระบวนการเริ่มต้น - metaphase บนโครโมโซมลูกสาว ด้วย Anaphase ของ mitosis ด้วยความช่วยเหลือของสปินเดิลพวกมันจะแยกออกจากขั้วที่ซึ่งนิวเคลียสทั้งสี่ถูกก่อตัวขึ้นและส่งผลให้เซลล์ haploid สี่เซลล์ ในเซลล์ของเนื้อเยื่อบางส่วนในระหว่างการพัฒนาของพวกเขาภายใต้อิทธิพลของปัจจัยบางอย่าง mitosis ไม่สมบูรณ์เกิดขึ้นและจำนวนของโครโมโซมในนิวเคลียสเป็นสองเท่าเนื่องจากความจริงที่ว่าพวกเขาไม่ได้แยกที่เสา เป็นผลมาจากการละเมิดธรรมชาติหรือเทียม, tetraploids และ polyploids เกิดขึ้น ด้วยความช่วยเหลือของไมโอซิสเซลล์สืบพันธุ์จะเกิดขึ้น - เซลล์สืบพันธุ์รวมถึงสปอร์องค์ประกอบของการสืบพันธุ์ทางเพศและการสืบพันธุ์แบบไม่อาศัยเพศของพืช (รูปที่ 18)
Amitosis เป็นฟิชชันโดยตรงของนิวเคลียส ด้วย amitosis แกนของการแบ่งจะไม่เกิดขึ้นและเปลือกของนิวเคลียสไม่สลายตัวเช่นเดียวกับในเซลล์ ก่อนหน้านี้ amitosis ถือเป็นรูปแบบดั้งเดิมของการแบ่ง ตอนนี้ได้รับการยอมรับว่ามีความเกี่ยวข้องกับการเสื่อมสภาพของร่างกาย มันเป็นเวอร์ชั่นที่เรียบง่ายของฟิชชันที่ซับซ้อนมากขึ้นของเคอร์เนล Amitosis เกิดขึ้นในเซลล์และเนื้อเยื่อของ nucellus, endosperm, tuber parenchyma, ก้านใบ, ฯลฯ
mitochondrion - เป็นอวัยวะที่มีลักษณะเป็นเกลียวกลมกลมยาวหรือแตกแขนง
แนวคิดของไมโทคอนเดรียถูกเสนอครั้งแรกโดยเบ็นด้าในปี 1897 มิโทคอนเดรียสามารถตรวจจับได้ในเซลล์ที่มีชีวิตโดยใช้คอนทราสต์ของเฟสและกล้องจุลทรรศน์แทรกแซงในรูปแบบของธัญพืชเม็ดหรือเส้นใย เหล่านี้เป็นโครงสร้างมือถือที่สามารถเคลื่อนย้ายผสานรวมกันได้ เมื่อย้อมด้วยวิธีพิเศษในเซลล์ที่ตายแล้วภายใต้กล้องจุลทรรศน์แบบแสงไมโทคอนเดรียจะมีลักษณะเป็นเม็ดเล็ก ๆ (เม็ด) ซึ่งกระจายอยู่ในไซโตพลาสซึมหรือกระจุกตัวในบางโซน
อันเป็นผลมาจากการทำลายของกลูโคสและไขมันต่อหน้าออกซิเจนพลังงานถูกสร้างขึ้นในไมโตคอนเดรียและสารอินทรีย์จะถูกเปลี่ยนเป็นน้ำและคาร์บอนไดออกไซด์ ด้วยวิธีนี้สิ่งมีชีวิตสัตว์ได้รับพลังงานพื้นฐานที่จำเป็นสำหรับชีวิต พลังงานถูกสะสมในอะดีโนซีนไตรฟอสเฟต (ATP) และแม่นยำยิ่งขึ้นในพันธบัตรมาโคร หน้าที่ของไมโตคอนเดรียนั้นเกี่ยวข้องกับการออกซิเดชั่นของสารประกอบอินทรีย์และการใช้พลังงานที่ปล่อยออกมาในระหว่างการสลายตัวของการสังเคราะห์โมเลกุล ATP ดังนั้นไมโตคอนเดรียจึงมักถูกเรียกว่าสถานีพลังงานของเซลล์หรือออร์แกเนลล์ของการหายใจของเซลล์ ATP ทำหน้าที่เป็นผู้จัดหาพลังงานถ่ายโอนหนึ่งในกลุ่มเทอร์มินัลฟอสเฟตที่อุดมด้วยพลังงานไปยังโมเลกุลอื่นและเปลี่ยนเป็น ADP
มีความเชื่อกันว่าในวิวัฒนาการไมโตคอนเดรียเป็นจุลินทรีย์โปรคาริโอติกที่กลายเป็น symbiotics ในร่างกายของเซลล์โบราณ ต่อจากนั้นพวกเขาจำเป็นอย่างยิ่งซึ่งเกี่ยวข้องกับการเพิ่มขึ้นของปริมาณออกซิเจนในชั้นบรรยากาศของโลก ในอีกด้านหนึ่งไมโตคอนเดรียได้กำจัดออกซิเจนพิษส่วนเกินสำหรับเซลล์และอีกด้านหนึ่งให้พลังงาน
หากปราศจากไมโตคอนเดรียเซลล์จะไม่สามารถใช้ออกซิเจนเป็นสารที่ให้พลังงานและสามารถตอบสนองความต้องการพลังงานของมันผ่านกระบวนการแบบไม่ใช้ออกซิเจน ดังนั้นออกซิเจนจึงเป็นพิษ แต่พิษมีความสำคัญต่อเซลล์และออกซิเจนส่วนเกินนั้นมีอันตรายเช่นเดียวกับการขาด
Mitochondria สามารถเปลี่ยนรูปร่างของพวกเขาและย้ายไปยังพื้นที่เหล่านั้นของเซลล์ที่ต้องการมากที่สุด ดังนั้นใน cardiomyocytes, ไมโตคอนเดรียอยู่ใกล้ myofibrils, ในเซลล์ของ tubules ไตใกล้กับ basal protrusions, ฯลฯ เซลล์ประกอบด้วยเซลล์ไมโทคอนเดรียถึงหนึ่งพันและจำนวนของมันขึ้นอยู่กับกิจกรรมของเซลล์
Mitochondria มีขนาดขวางเฉลี่ย 0.5 ... 3 ไมครอน ไมโตคอนเดรียขนาดเล็กกลางใหญ่และใหญ่ขึ้นอยู่กับขนาด (พวกมันก่อตัวเป็นเครือข่ายแขนง - ไมโทคอนเดรีย reticulum) ขนาดและจำนวนของไมโตคอนเดรียนั้นสัมพันธ์กับกิจกรรมของเซลล์และการใช้พลังงาน พวกมันแปรปรวนอย่างมากและขึ้นอยู่กับกิจกรรมของเซลล์ปริมาณออกซิเจนอิทธิพลของฮอร์โมนสามารถบวมเปลี่ยนจำนวนและโครงสร้างของคริสเตแตกต่างกันไปในจำนวนรูปร่างและขนาดเช่นเดียวกับกิจกรรมของเอนไซม์
ความหนาแน่นรวมของไมโตคอนเดรียระดับการพัฒนาของพื้นผิวด้านในและตัวบ่งชี้อื่น ๆ ขึ้นอยู่กับความต้องการพลังงานของเซลล์ ในเซลล์เม็ดเลือดขาวมีไมโตคอนเดรียเพียงไม่กี่ตัวและในเซลล์ตับมี 2 ... 3 พัน
Mitochondria ประกอบด้วยเมทริกซ์, เยื่อหุ้มชั้นใน, พื้นที่ perimitochondrial และเยื่อหุ้มชั้นนอก เยื่อหุ้มเซลล์ยลไมโตคอนเดรียด้านนอกจะแยกออร์แกเนลล์ออกจากไฮยาโลพลาสซึม โดยปกติจะมีรูปทรงที่เรียบและถูกปิดเพื่อให้เป็นถุงเมมเบรน
เยื่อหุ้มชั้นนอกจากด้านในจะถูกคั่นด้วยพื้นที่ perimitochondrial ที่มีความกว้างประมาณ 10 ... 20 นาโนเมตร เมมเบรนภายในยล จำกัด เนื้อหาภายในที่แท้จริงของไมโตคอนเดรีย - เมทริกซ์ เยื่อหุ้มชั้นในนั้นยื่นออกมามากมายภายในไมโตคอนเดรียซึ่งมีลักษณะเป็นสันเขาแบนหรือคริสเต
ในรูปร่าง cristae สามารถอยู่ในรูปแบบของแผ่น (trabecular) และ tubules (หลากหลายที่ตัด) และพวกเขาจะถูกนำไปตามยาวหรือตามขวางด้วยความเคารพไมโตคอนเดรีย
ไมโตคอนเดรียแต่ละตัวเต็มไปด้วยเมทริกซ์ซึ่งใน micrographs อิเล็กตรอนมีลักษณะหนาแน่นกว่าไซโตพลาสซึมโดยรอบ mitochondrial matrix นั้นเป็นเนื้อเดียวกัน (homogeneous) ซึ่งบางครั้งมีความละเอียดของความหนาแน่นของอิเล็กตรอนหลายชนิด มันเผยให้เห็นเส้นใยบาง ๆ ที่มีความหนาประมาณ 2 ... 3 นาโนเมตรและเม็ดที่มีขนาดประมาณ 15 ... 20 นาโนเมตร เมทริกซ์เส้นเป็นโมเลกุล DNA และเม็ดเล็ก ๆ เป็นไมโตคอนเดรียไรโบโซม เมทริกซ์นั้นมีเอ็นไซม์, DNA แบบวนรอบเดียว, ไซโคลอิสระ, ไรโบโซมแบบไมโตคอนเดรีย, Ca 2+ ไอออนจำนวนมาก
ระบบอิสระของการสังเคราะห์โปรตีนของไมโทคอนเดรียนั้นแสดงโดยโมเลกุลดีเอ็นเอที่ปราศจากฮิสโตน ดีเอ็นเอสั้นมีรูปร่างของวงแหวน (วงจร) และมี 37 ยีน ซึ่งแตกต่างจาก DNA นิวเคลียร์มีจริงไม่มีลำดับนิวคลีโอไทด์ที่ไม่ใช่การเข้ารหัสในนั้น คุณสมบัติของโครงสร้างและองค์กรนำ DNA ของ mitochondrial มารวมกันกับ DNA ของเซลล์แบคทีเรีย ใน DNA ของไมโตคอนเดรียการสังเคราะห์โมเลกุลอาร์เอ็นเอของประเภทต่างๆเกิดขึ้น: ข้อมูลการถ่ายโอน (การขนส่ง) และไรโบโซม ข้อมูล RNA ของไมโทคอนเดรียไม่ได้ถูกประกบกัน (ส่วนตัดที่ไม่มีการโหลดข้อมูล) โมเลกุลของไมโตคอนเดรียขนาดเล็กไม่สามารถตรวจสอบการสังเคราะห์โปรตีนไมโตคอนเดรียทั้งหมดได้ โปรตีนไมโตคอนเดรียส่วนใหญ่อยู่ภายใต้การควบคุมทางพันธุกรรมของนิวเคลียสของเซลล์และถูกสังเคราะห์ในไซโตพลาสซึมเนื่องจาก DNA ของไมโทคอนเดรียมีการแสดงออกอย่างอ่อนและสามารถให้การก่อตัวของเอนไซม์โซ่ออกซิเดชัน DNA ของไมโตคอนเดรียไม่ได้เข้ารหัสโปรตีนที่อยู่ในเยื่อบุและมีโครงสร้างโปรตีนที่รับผิดชอบการรวมกันของคอมเพล็กซ์โปรตีนที่ใช้งานได้ของเยื่อหุ้มไมโตคอนเดรียที่เหมาะสม โปรตีนที่ทำหน้าที่ขนส่งก็ถูกสังเคราะห์เช่นกัน ระบบการสังเคราะห์โปรตีนดังกล่าวไม่ได้ให้การทำงานทั้งหมดของไมโตคอนเดรียดังนั้นเอกสิทธิ์ของไมโตคอนเดรียจึง จำกัด และสัมพันธ์กัน
ในสัตว์เลี้ยงลูกด้วยนม, ไมโทคอนเดรียในระหว่างการปฏิสนธิจะถูกส่งผ่านทางไข่เท่านั้นและสเปิร์มจะนำดีเอ็นเอนิวเคลียสเข้าสู่สิ่งมีชีวิตใหม่
ในเมทริกซ์ของไมโทคอนเดรียไรโบโซมเกิดขึ้นที่แตกต่างจากไรโซไซโตพลาสซึม พวกเขามีส่วนร่วมในการสังเคราะห์โปรตีนไมโตคอนเดรียจำนวนหนึ่งซึ่งไม่ได้เข้ารหัสโดยนิวเคลียส ไรโบโซมระดับยลมีจำนวนการตกตะกอน 60 (ไม่เหมือนกับไซโตพลาสซึมที่มีจำนวนการตกตะกอน 80) หมายเลขการตกตะกอนคืออัตราการสะสมในระหว่างการปั่นแยกและหมุนเหวี่ยงเป็นพิเศษ ในโครงสร้างไมโตคอนเดรียไรโบโซมอยู่ใกล้ไรโบโซมของสิ่งมีชีวิตที่เป็น prokaryotic แต่มีขนาดเล็กกว่าและไวต่อยาปฏิชีวนะบางชนิด (chloramphenicol, tetracycline ฯลฯ )
เยื่อหุ้มไมโตคอนเดรียด้านในมีระดับหัวกะทิสูงในการขนส่งสาร เอนไซม์ของโซ่ออกซิเดชัน phosphorylation โปรตีนพาหะอิเล็กตรอน ATP, ADP, ไพรูเวตและระบบการขนส่งอื่น ๆ ที่อยู่ติดกันใกล้ชิดกันติดอยู่กับพื้นผิวด้านในอันเป็นผลมาจากการจัดเรียงของเอนไซม์ในเยื่อหุ้มชั้นในคอนจูเกชั่น ประสิทธิภาพของกระบวนการเร่งปฏิกิริยา
กล้องจุลทรรศน์อิเล็กตรอนเผยให้เห็นอนุภาคของเห็ดที่ยื่นออกมาในรูของเมทริกซ์ พวกเขามีกิจกรรม ATP- สังเคราะห์ (ฟอร์ม ATP จาก ADP) การขนส่งอิเล็คตรอนเกิดขึ้นตามสายโซ่การหายใจซึ่งมีการแปลเป็นภาษาท้องถิ่นในเยื่อหุ้มชั้นในซึ่งมีเอ็นไซม์ขนาดใหญ่สี่คอมเพกต์ เมื่ออิเล็กตรอนผ่านห่วงโซ่การหายใจไอออนของไฮโดรเจนจะถูกสูบออกจากเมทริกซ์ไปยังพื้นที่ perimitochondrial ซึ่งช่วยให้แน่ใจว่าการก่อตัวของการไล่ระดับโปรตอน (ปั๊ม) พลังงานของการไล่ระดับสีนี้ (ความแตกต่างในความเข้มข้นของสารและการก่อตัวของศักยภาพเยื่อหุ้มเซลล์) ใช้สำหรับการสังเคราะห์ ATP และการขนส่งเมตาบอไลต์และไอออนอนินทรีย์ โปรตีนพาหะที่มีอยู่ในเมมเบรนด้านในจะลำเลียงสารอินทรีย์ฟอสเฟต ATP, ADP, กรดอะมิโน, กรดไขมัน, กรดไตรและกรดคาร์บอกซิลิกผ่าน
เยื่อหุ้ม mitochondrial ด้านนอกนั้นสามารถซึมผ่านไปยังสารที่มีน้ำหนักโมเลกุลต่ำได้มากกว่าเนื่องจากมีช่องทางโปรตีนไฮโดรฟิลิกหลายช่องทาง คอมเพล็กซ์ตัวรับเฉพาะตั้งอยู่บนเยื่อหุ้มชั้นนอกซึ่งโปรตีนจากเมทริกซ์จะถูกส่งไปยังพื้นที่รอบนอก
ในแง่ขององค์ประกอบทางเคมีและคุณสมบัติเมมเบรนด้านนอกอยู่ใกล้กับเยื่อหุ้มเซลล์อื่นและพลาสโมเลมา มันมีเอนไซม์ที่เผาผลาญไขมันเปิดใช้งาน (เร่งปฏิกิริยา) การแปลงของเอมีนอะมิโนออกซิเดส หากเอนไซม์ของเยื่อหุ้มชั้นนอกยังคงทำงานอยู่สิ่งนี้จะเป็นตัวบ่งชี้ความปลอดภัยในการทำงานของไมโตคอนเดรีย
มีสองส่วนย่อยอิสระในไมโทคอนเดรีย เนื่องจากพื้นที่ perimitochondrial หรือห้อง mitochondrial ด้านนอก (sub-compartment ภายนอก) เกิดขึ้นเนื่องจากการแทรกซึมของโปรตีนเชิงซ้อนของ hyaloplasm ทำให้ sub-compartment ภายใน (เมทริกซ์ยล) นั้นเกิดขึ้นบางส่วนเนื่องจากกิจกรรมสังเคราะห์ของยลยลดีเอ็นเอ sub-compartment ภายใน (เมทริกซ์) มี DNA, RNA และไรโบโซม มันมีไอออน Ca 2+ ในระดับสูงเมื่อเปรียบเทียบกับ hyaloplasm ในส่วนย่อยภายนอกไฮโดรเจนไอออนสะสม กิจกรรมของเอนไซม์ขององค์ประกอบย่อยภายนอกและภายในองค์ประกอบของโปรตีนจะแตกต่างกันมาก ช่องย่อยภายในมีความหนาแน่นของอิเล็กตรอนสูงกว่าภายนอก
เครื่องหมายไมโตคอนเดรียพิเศษคือไซโตโครมออกซิเดสและเอนไซม์ดีไฮโดรจีเนสซึ่งช่วยในการตรวจสอบคุณสมบัติเชิงปริมาณของกระบวนการพลังงานในไมโตคอนเดรีย
หน้าที่หลักของไมโตคอนเดรีย - การสังเคราะห์ ATP เริ่มแรกน้ำตาล (กลูโคส) จะถูกทำลายใน hyaloplasm ไปยังกรดแลคติกและไพรูริก (ไพรูเวต) ด้วยการสังเคราะห์ ATP จำนวนเล็กน้อยพร้อมกัน เป็นผลมาจาก glycolysis ของกลูโคสหนึ่งโมเลกุลจะใช้ ATP สองโมเลกุลและสี่รูปแบบ ดังนั้นเพียงสองโมเลกุล ATP ประกอบขึ้นเป็นดุลยภาพ กระบวนการเหล่านี้เกิดขึ้นโดยไม่มีออกซิเจน (anaerobic glycolysis)
การผลิตพลังงานในระยะต่อมาทั้งหมดเกิดขึ้นในกระบวนการออกซิเดชั่นแอโรบิกซึ่งเป็นการสังเคราะห์ ATP จำนวนมาก ในกรณีนี้สารอินทรีย์จะถูกทำลายไปยัง CO 2 และน้ำ ออกซิเดชันจะมาพร้อมกับการส่งโปรตอนไปยังผู้ตอบรับ ปฏิกิริยาเหล่านี้ดำเนินการโดยใช้เอ็นไซม์จำนวนหนึ่งของวัฏจักรกรดไตรคาร์บอกซิลิกซึ่งอยู่ในเมทริกซ์ไมโทคอนเดรีย
ระบบการถ่ายโอนอิเล็กตรอนและ phosphorylation ที่เกี่ยวข้องของ ADP (oxidative phosphorylation) ถูกสร้างขึ้นในเยื่อหุ้มคริสตัล ในกรณีนี้การถ่ายโอนอิเล็กตรอนจากโปรตีนตัวรับอิเล็กตรอนหนึ่งไปยังอีกเกิดขึ้นและในที่สุดความผูกพันของพวกเขากับออกซิเจนเป็นผลมาจากการที่น้ำจะเกิดขึ้น ในเวลาเดียวกันส่วนหนึ่งของพลังงานที่ปล่อยออกมาในระหว่างการเกิดออกซิเดชันในห่วงโซ่การถ่ายโอนอิเล็กตรอนจะถูกเก็บไว้ในรูปแบบของพันธะแบบมหภาคระหว่าง ADF phosphorylation ซึ่งจะนำไปสู่การก่อตัวของโมเลกุล ATP จำนวนมาก - พลังงานภายในเซลล์หลัก กระบวนการเกิดออกซิเดชันของฟอสโฟรีเลชันเกิดขึ้นที่เยื่อหุ้มของไมโทคอนเดรียคริสทีโดยใช้โปรตีนสายโซ่ออกซิเดชั่นและเอนไซม์ ADP ATP synthetase phosphorylation ADP ATP เป็นผลมาจาก oxidative phosphorylation โมเลกุล 36 ATP เกิดจากกลูโคสโมเลกุลหนึ่ง
สำหรับฮอร์โมนและสารบางชนิดในไมโตคอนเดรียมีตัวรับ (ความสัมพันธ์) เฉพาะ Triiodothyronine เร่งการสังเคราะห์ของไมโตคอนเดรีย Interleukin-1 และความเข้มข้นสูงของ triiodothyronine uncouple phosphorylation ออกซิเดชันโซ่ทำให้เกิดอาการบวมยลซึ่งมาพร้อมกับการเพิ่มขึ้นของการก่อตัวของพลังงานความร้อน
ไมโตคอนเดรียใหม่เกิดขึ้นจากการแบ่งการหดหรือการแตกหน่อ ในกรณีหลังมีการสร้างโปรโตมิโตคอนเดรียเพิ่มขึ้นเรื่อย ๆ ในขนาด
Protomitochondria เป็น organelle ขนาดเล็กที่มีเยื่อหุ้มชั้นนอกและชั้นใน เยื่อหุ้มชั้นในไม่ได้มีหรือมีคริสตี้ที่ด้อยพัฒนา ออร์แกเนลล่ามีลักษณะเป็นระดับฟอสฟอรัสในเลือดต่ำ ด้วยการก่อตัวของการหดตัวเนื้อหาของ mitochondria มีการกระจายระหว่างสอง organelles ใหม่ค่อนข้างใหญ่ ด้วยวิธีการใด ๆ ของการสืบพันธุ์ไมโตคอนเดรียที่เพิ่งสร้างใหม่แต่ละตัวมีจีโนมของตัวเอง
ไมโตคอนเดรียที่เก่ากว่านั้นถูกทำลายโดย autolysis (เซลล์ย่อยสลายด้วยตนเองโดยใช้ lysosomes) เพื่อสร้าง autolysosomes จาก autolysosome ร่างกายที่เหลือจะเกิดขึ้น ด้วยการย่อยที่สมบูรณ์เนื้อหาของร่างกายที่เหลือซึ่งประกอบด้วยสารอินทรีย์ที่มีน้ำหนักโมเลกุลต่ำจะถูกขับออกมาโดย exocytosis ด้วยการย่อยที่ไม่สมบูรณ์ส่วนที่เหลือของไมโทคอนเดรียสามารถสะสมในเซลล์ในรูปแบบของชั้นหรือแกรนูลที่มี nipofuscin เกลือแคลเซียมที่ไม่ละลายน้ำสะสมในส่วนของไมโทคอนเดรียพร้อมกับการก่อตัวของผลึก - กลายเป็นปูน การสะสมของผลิตภัณฑ์ความเสื่อมยลยลสามารถนำไปสู่เซลล์เสื่อม
โครงสร้างและหน้าที่ของไมโตคอนเดรียเป็นปัญหาที่ค่อนข้างซับซ้อน การปรากฏตัวของออร์แกเนลล์เป็นลักษณะของสิ่งมีชีวิตนิวเคลียร์เกือบทั้งหมด - ทั้งสำหรับออโตโทรฟ (พืชที่สามารถสังเคราะห์แสง) และสำหรับเฮเทอโรโทรฟซึ่งเป็นสัตว์เกือบทุกชนิดพืชและเชื้อราบางชนิด
จุดประสงค์หลักของไมโทคอนเดรียคือการออกซิเดชั่นของสารอินทรีย์และการใช้พลังงานที่ปล่อยออกมาจากกระบวนการนี้ ด้วยเหตุนี้ออร์แกเนลล์จึงมีชื่อสถานีพลังงานเซลล์ที่สอง (ไม่เป็นทางการ) บางครั้งพวกเขาเรียกว่า "catabolism plastids"
ไมโตคอนเดรียคืออะไร
คำนี้มีต้นกำเนิดมาจากภาษากรีก แปลคำนี้หมายถึง "thread" (mitos), "seed" (chondrion) Mitochondria เป็น organoids ถาวรที่มีความสำคัญอย่างยิ่งสำหรับการทำงานปกติของเซลล์และทำให้เป็นไปได้สำหรับสิ่งมีชีวิตทั้งหมด

"สถานี" มีโครงสร้างภายในที่เฉพาะเจาะจงซึ่งแตกต่างกันไปขึ้นอยู่กับสถานะการทำงานของไมโตคอนเดรีย รูปร่างของพวกเขาสามารถเป็นสองประเภท - รูปไข่หรือเป็นรูปสี่เหลี่ยมผืนผ้า หลังมักจะมีลักษณะแตกแขนง จำนวนของ organelles ในหนึ่งเซลล์มีตั้งแต่ 150 ถึง 1500
กรณีพิเศษคือเซลล์สืบพันธุ์ มีออร์แกนิกเกลียวเพียงอันเดียวในสเปิร์มในขณะที่เซลล์สืบพันธุ์เพศหญิงมีไมโตคอนเดรียนับแสน ในเซลล์ organelles ไม่ได้รับการแก้ไขในที่เดียว แต่สามารถเคลื่อนที่ไปตามไซโตพลาสซึมรวมเข้าด้วยกัน ขนาดของพวกเขาคือ 0.5 ไมครอนความยาวสามารถเข้าถึง 60 ไมครอนในขณะที่ตัวเลขขั้นต่ำคือ 7 ไมครอน

การกำหนดขนาดของ "สถานีพลังงาน" ไม่ใช่เรื่องง่าย ความจริงก็คือเมื่อตรวจสอบด้วยกล้องจุลทรรศน์อิเล็กตรอนมีเพียงบางส่วนของออร์แกเนลล์เท่านั้นที่เข้าไปในส่วน มันเกิดขึ้นได้ว่าไมโทคอนเดรียเป็นเกลียวมีหลายส่วนที่สามารถแยกออกเป็นโครงสร้างอิสระ
มีเพียงภาพสามมิติเท่านั้นที่จะช่วยให้เราค้นพบโครงสร้างเซลลูลาร์ที่แน่นอนและเข้าใจว่าเรากำลังพูดถึงออร์การอยด์แยกกัน 2-5 หรือประมาณหนึ่งอันด้วยไมโตคอนเดรียที่ซับซ้อน
คุณสมบัติโครงสร้าง
เยื่อหุ้มเซลล์ยลประกอบด้วยสองชั้น: ภายนอกและภายใน หลังรวมถึงผลพลอยได้ต่างๆและเท่าที่มีรูปร่างเหมือนใบไม้และท่อ
เมมเบรนแต่ละตัวมีองค์ประกอบทางเคมีพิเศษเอนไซม์บางชนิดและวัตถุประสงค์เฉพาะ เปลือกนอกจากด้านในคั่นด้วยช่องว่างระหว่างกันด้วยความหนา 10-20 นาโนเมตร
โครงสร้างของออร์แกเนลล์ในรูปพร้อมคำบรรยายมีความชัดเจนมาก

โครงสร้างของไมโตคอนเดรีย
โดยการดูที่แผนภาพโครงสร้างคำอธิบายต่อไปนี้สามารถทำได้ พื้นที่ความหนืดภายในไมโตคอนเดรียเรียกว่าเมทริกซ์ องค์ประกอบของมันสร้างสภาพแวดล้อมที่เอื้ออำนวยต่อการไหลของกระบวนการทางเคมีที่จำเป็น มันประกอบด้วยเม็ดกล้องจุลทรรศน์ที่ส่งเสริมปฏิกิริยาและกระบวนการทางชีวเคมี (ตัวอย่างเช่นพวกเขาสะสมไอออนไกลโคเจนและสารอื่น ๆ )
เมทริกซ์ประกอบด้วย DNA, โคเอนไซม์, ไรโบโซม, t-RNA, ไอออนอนินทรีย์ ATP synthase และ cytochromes ตั้งอยู่บนพื้นผิวของชั้นในของเปลือก เอนไซม์มีส่วนร่วมในกระบวนการต่าง ๆ เช่นวงจร Krebs (CTK), ออกซิเดชั่นฟอสโฟรีเลชัน ฯลฯ
ดังนั้นหน้าที่หลักของ organoid จึงดำเนินการโดยทั้ง matrix และด้านในของเปลือก
ฟังก์ชั่นยล
วัตถุประสงค์ของ "โรงไฟฟ้า" สามารถจำแนกได้สองงานหลัก:
- การผลิตพลังงาน: กระบวนการออกซิเดชั่นถูกดำเนินการในตัวพวกมันด้วยการปล่อยโมเลกุล ATP ในภายหลัง
- การเก็บข้อมูลทางพันธุกรรม
- การมีส่วนร่วมในการสังเคราะห์ฮอร์โมนกรดอะมิโนและโครงสร้างอื่น ๆ
กระบวนการออกซิเดชันและการผลิตพลังงานเกิดขึ้นในหลายขั้นตอน:


ภาพวาดแผนผังของการสังเคราะห์ ATP
มันเป็นที่น่าสังเกตว่า: อันเป็นผลมาจากวงจร Krebs (รอบกรดซิตริก) โมเลกุล ATP จะไม่เกิดขึ้นโมเลกุลจะถูกออกซิไดซ์และปล่อยก๊าซคาร์บอนไดออกไซด์ นี่เป็นขั้นตอนกลางระหว่าง glycolysis และห่วงโซ่การขนส่งอิเล็กตรอน
ตาราง "ฟังก์ชั่นและโครงสร้างของไมโทคอนเดรีย"

สิ่งที่กำหนดจำนวนไมโตคอนเดรียในเซลล์
จำนวนของสารอินทรีย์ที่สะสมอยู่ใกล้กับส่วนต่าง ๆ ของเซลล์ที่มีความต้องการทรัพยากรพลังงาน โดยเฉพาะอย่างยิ่งจะมีการรวบรวมออร์แกเนลล์จำนวนมากในบริเวณ myofibrils ซึ่งเป็นส่วนหนึ่งของเซลล์กล้ามเนื้อที่รับประกันการหดตัว

ในเซลล์สืบพันธุ์เพศชายโครงสร้างจะถูก จำกัด วงรอบแกนแฟลเจลลัม - ซึ่งสันนิษฐานว่าความต้องการ ATP นั้นเกิดจากการเคลื่อนที่อย่างคงที่ของหางของเซลล์สืบพันธุ์ การจัดเรียงของไมโทคอนเดรียในโปรโตซัวซึ่งใช้ cilia พิเศษสำหรับการเคลื่อนไหวดูเหมือนว่า organelles สะสมอยู่ภายใต้เมมเบรนที่ฐานของพวกเขาในลักษณะเดียวกัน
สำหรับเซลล์ประสาทพบว่ามีการแปลของไมโตคอนเดรียใกล้กับซินไซด์ซึ่งสัญญาณของระบบประสาทถูกส่งผ่าน ในการสังเคราะห์โปรตีนในเซลล์ organelles จะสะสมในโซน ergastoplasm ซึ่งเป็นแหล่งพลังงานที่ให้กระบวนการนี้
ผู้ค้นพบไมโตคอนเดรีย
โครงสร้างของเซลล์ได้รับชื่อในปี 1897-1898 จาก C. Brand Otto Wagburg สามารถพิสูจน์การเชื่อมต่อของการหายใจของเซลล์กับ mitochondria ในปี 1920
ข้อสรุป
Mitochondria เป็นส่วนประกอบที่สำคัญที่สุดของเซลล์สิ่งมีชีวิตทำหน้าที่เป็นสถานีพลังงานที่ผลิตโมเลกุล ATP ดังนั้นจึงมั่นใจได้ว่ากระบวนการของกิจกรรมที่สำคัญของเซลล์
การทำงานของไมโตคอนเดรียขึ้นอยู่กับการเกิดออกซิเดชันของสารประกอบอินทรีย์ทำให้เกิดพลังงานศักย์
เยื่อหุ้มชั้นนอก
เยื่อหุ้มชั้นใน
มดลูก m-on, เมทริกซ์, crista. มันมีรูปทรงที่เรียบไม่ก่อรูปแบบยื่นออกมาหรือเท่า มันคิดเป็นประมาณ 7% ของพื้นที่ของเยื่อหุ้มเซลล์ทั้งหมด ความหนาของมันอยู่ที่ประมาณ 7 นาโนเมตรมันไม่ได้เชื่อมต่อกับเยื่ออื่น ๆ ของไซโตพลาสซึมและปิดตัวเองดังนั้นมันจึงเป็นถุงเมมเบรน เยื่อหุ้มชั้นนอกแยกออกจากด้านใน อวกาศ intermembrane กว้างประมาณ 10-20 นาโนเมตร เยื่อหุ้มชั้นใน (หนาประมาณ 7 นาโนเมตร) จำกัด ขอบเขตเนื้อหาภายในที่แท้จริงของไมโตคอนเดรีย
เมทริกซ์หรือ mitoplasm ของมัน คุณลักษณะเฉพาะของเยื่อหุ้มชั้นในของไมโทคอนเดรียคือความสามารถในการก่อส่วนยื่นออกจำนวนมากภายในไมโตคอนเดรีย ส่วนที่ยื่นออกมาดังกล่าวส่วนใหญ่มักจะมีรูปแบบของแนวสันเขาหรือคริส ระยะห่างระหว่างเยื่อหุ้มเซลล์ใน Crista อยู่ที่ประมาณ 10-20 นาโนเมตร คริสเตมักจะแตกแขนงหรือก่อตัวเป็นรูปนิ้วกระบวนการงอและไม่มีทิศทางที่เด่นชัด ในโปรโตซัวสาหร่ายที่มีเซลล์เดียวในบางเซลล์ของพืชและสัตว์ที่สูงกว่าผลพลอยได้ของเยื่อหุ้มชั้นในจะมีลักษณะคล้ายท่อ (cristae ท่อ)
ไมโตคอนเดรียเมทริกซ์มีโครงสร้างที่เป็นเนื้อเดียวกันอย่างละเอียดบางครั้งบาง ๆ เส้นใยที่เก็บรวบรวมในลูกบอล (ประมาณ 2-3 นาโนเมตร) และตรวจพบเม็ดเล็ก ๆ ประมาณ 15-20 นาโนเมตร ตอนนี้มันกลายเป็นที่รู้จักกันว่าเส้นเมทริกซ์ยลเป็นโมเลกุลดีเอ็นเอในการประกอบของนิวทรอยด์ยลและเม็ดเล็ก ๆ เป็นยลไรโบโซม
ฟังก์ชั่นยล
1. ในการสังเคราะห์เอทีพี mitochondria เกิดขึ้น (ดูออกซิเดชัน phosphorylation)
ค่า Intermembrane pH ~ 4, เมทริกซ์ค่า pH ~ 8 | ปริมาณโปรตีนใน m: 67% เป็นเมทริกซ์, 21% เป็นการค้นพบของฉัน, 6% เป็นด้านในของฉันและ 6% อยู่ในเหมือง
Handrioma - ระบบยลเดียว
m-na ด้านนอก: porin-pores อนุญาตให้ผ่านได้สูงสุด 5 kD | Inner m-na: cardiolipin-impervious m-well สำหรับไอออน |
การผลิตระหว่างประเทศ: กลุ่มเอนไซม์ phosphorylate นิวคลีโอไทด์และนิวคลีโอไทด์น้ำตาล
เมตรภายในเมื่อ:
เมทริกซ์: เอนไซม์เมตาบอลิซึม - ออกซิเดชันของไขมัน, ออกซิเดชั่นของคาร์โบไฮเดรต, วัฏจักรกรด tricarboxylic, วงจร Krebs
ต้นกำเนิดมาจากแบคทีเรีย: อะมีบา Pelomyxa palustris ไม่มีเอกภาพจากยูคาริโอตอาศัยอยู่ใน symbiosis กับแบคทีเรียแอโรบิก | DNA ของตัวเอง แบคทีเรียคล้าย OX
DNA ยล
การแบ่ง Myochondria
การจำลองแบบ
ในเฟส การจำลองแบบไม่เกี่ยวข้องกับ S-phase | ในระหว่างรอบ cl, mitochis จะถูกแบ่งออกเป็นสองครั้งครั้งเดียว, สร้างการรัด, การหดตัวครั้งแรกใน m- ไม่ | ~ 16.5 kb | แหวนเข้ารหัส 2 rRNA 22 tRNA และโปรตีน 13 ตัว
การขนส่งโปรตีน: สัญญาณเปปไทด์ | amphiphilic curl | ตัวรับรู้ยล
ฟอสฟอรัสออกซิเดชัน
วงจรการถ่ายโอนอิเล็กตรอน
ATP synthase
ในเซลล์ตับ m มีชีวิตอยู่ ~ 20 วันแบ่งยลโดยการก่อตัวของการหดตัว
16569 bp \u003d 13 โปรตีน, 22tRNA, 2pRNA | m-na ด้านนอกเรียบ (porins - การซึมผ่านของโปรตีนสูงถึง 10 kDa) พับด้านใน (crista) m-na (โปรตีน 75%): โปรตีนตัวพาขนส่ง, f-you, ส่วนประกอบของห่วงโซ่ทางเดินหายใจและ ATP synthase, cardiolipin) เมทริกซ์ ( ผสานกับวงจร f-tami citrate) inter 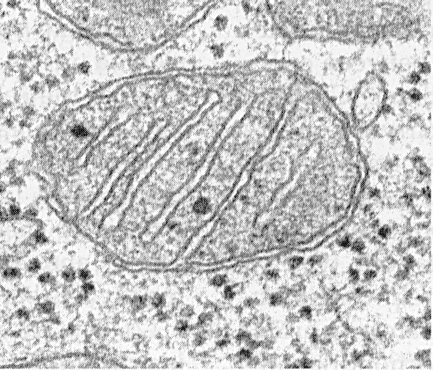
Mitochondria หรือ chondriosomes (จากกรีก. Mitos - ด้าย, chondrion - เมล็ดพืช, soma - body) เป็น organelles เม็ดหรือเส้นใยอยู่ในพลาสซึมของโปรโตซัว, พืชและสัตว์. ไมโตคอนเดรียสามารถสังเกตได้ในเซลล์ที่มีชีวิตเนื่องจากมีความหนาแน่นค่อนข้างสูง ในเซลล์ที่มีชีวิตไมโตคอนเดรียสามารถเคลื่อนย้ายขยับรวมเข้าด้วยกัน
ในสายพันธุ์ที่แตกต่างกันขนาดของไมโตคอนเดรียนั้นแปรปรวนมากและรูปร่างของมันก็แปรผัน (รูปที่ 199) อย่างไรก็ตามในเซลล์ส่วนใหญ่ความหนาของโครงสร้างเหล่านี้ค่อนข้างคงที่ (ประมาณ 0.5 μm) และความยาวมีความผันผวนสูงถึง 7-60 ไมครอนในรูปแบบไส้
การศึกษาขนาดและจำนวนไมโตคอนเดรียไม่ใช่เรื่องง่าย นี่เป็นเพราะความจริงที่ว่าขนาดและจำนวนของไมโตคอนเดรียที่มองเห็นได้ในบางเฉียบไม่ตรงกับความเป็นจริง
การคำนวณแบบธรรมดาแสดงให้เห็นว่ามี 200 ไมโตคอนเดรียอยู่ในเซลล์ตับ ซึ่งแสดงถึงมากกว่า 20% ของพลาสซึมทั้งหมดและประมาณ 30-35% ของปริมาณโปรตีนทั้งหมดในเซลล์ พื้นที่ผิวของไมโตคอนเดรียทั้งหมดของเซลล์ตับนั้นมีขนาดใหญ่กว่าผิวของพลาสมาเมมเบรนประมาณ 4-5 เท่า ไมโตคอนเดรียส่วนใหญ่อยู่ในไข่ (ประมาณ 300,000) และในอะมีบาอลหม่านยักษ์ (สูงถึง 500,000)
ในเซลล์พืชสีเขียวจำนวนไมโตคอนเดรียน้อยกว่าในเซลล์สัตว์เนื่องจากคลอโรพลาสต์สามารถทำหน้าที่เป็นส่วนหนึ่งของมันได้
การแปลของไมโทคอนเดรียในเซลล์นั้นแตกต่างกัน โดยทั่วไปแล้วไมโทคอนเดรียจะสะสมอยู่ใกล้กับบริเวณของไซโตพลาสซึมซึ่งมีความต้องการ ATP ที่เกิดขึ้นในไมโตคอนเดรีย ดังนั้นในกล้ามเนื้อโครงร่างไมโตคอนเดรียจึงอยู่ใกล้ myofibrils ในสเปิร์มไมโตคอนเดรียก่อตัวเป็นเกลียวรอบแกนแฟลเจลลัม นี่อาจเป็นเพราะความต้องการใช้ ATP เพื่อย้ายหางของอสุจิ ในทำนองเดียวกันในโปรโตซัวและในเซลล์อื่น ๆ ที่มี cilia mitochondria จะถูกแปลเป็นภาษาท้องถิ่นโดยตรงภายใต้เยื่อหุ้มเซลล์ที่ฐานของ cilia ซึ่งต้องใช้ ATP ในการทำงาน ในแอกซอนของเซลล์ประสาทไมโตคอนเดรียอยู่ใกล้กับซินเซสซึ่งกระบวนการส่งผ่านแรงกระตุ้นเส้นประสาทเกิดขึ้น ในเซลล์หลั่งที่สังเคราะห์โปรตีนจำนวนมากไมโตคอนเดรียสัมพันธ์กับโซน ergastoplasma อย่างใกล้ชิด พวกเขาอาจให้ ATP เพื่อเปิดใช้งานกรดอะมิโนและการสังเคราะห์โปรตีนในไรโบโซม
โครงสร้างพื้นฐานของไมโตคอนเดรีย
Mitochondria ไม่ว่าจะมีขนาดหรือรูปร่างใดก็ตามมีโครงสร้างที่เป็นสากล Mitochondria นั้น จำกัด อยู่เพียงสองแผ่นเท่านั้น (รูปที่ 205) เยื่อหุ้มเซลล์ยลไมโตคอนเดรียชั้นนอกแยกออกจากไฮยาโลพลาสซึมมีรูปร่างที่ราบเรียบไม่ก่อรูปหรือยื่นออกมาและมีความหนาประมาณ 7 นาโนเมตร มันคิดเป็นประมาณ 7% ของพื้นที่ของเยื่อหุ้มเซลล์ทั้งหมด เมมเบรนไม่ได้เชื่อมต่อกับเยื่อหุ้มเซลล์อื่นของไซโตพลาสซึมปิดตัวเองและเป็นถุงเมมเบรน เยื่อหุ้มชั้นนอกจากด้านในจะถูกคั่นด้วยช่องว่างระหว่างกันด้วยความกว้างประมาณ 10-20 นาโนเมตร เยื่อหุ้มชั้นใน (ประมาณ 7 นาโนเมตรหนา) จำกัด เนื้อหาภายในที่แท้จริงของไมโตคอนเดรียเมทริกซ์หรือไมโทพลาสซึม เยื่อหุ้มไมโตคอนเดรียชั้นในนั้นก่อตัวเป็นนูนจำนวนมากภายในไมโตคอนเดรีย ส่วนที่ยื่นออกมาดังกล่าวส่วนใหญ่มักจะมีรูปแบบของแนวสันเขาหรือคริส
พื้นผิวทั้งหมดของเยื่อหุ้มเซลล์ยลภายในในเซลล์ตับจะอยู่ที่ประมาณหนึ่งในสามของพื้นผิวของเยื่อหุ้มเซลล์ทั้งหมด ไมโตคอนเดรียของเซลล์กล้ามเนื้อหัวใจมีคริสเตจำนวนมากเป็นสามเท่าของตับไมโตคอนเดรียซึ่งสะท้อนถึงความแตกต่างในการทำงานของไมโตคอนเดรียของเซลล์ต่าง ๆ ระยะห่างระหว่างเยื่อหุ้มเซลล์ใน Crista อยู่ที่ประมาณ 10-20 นาโนเมตร
คริสทีไมโตคอนเดรียลที่ยื่นออกมาจากเยื่อหุ้มชั้นในและขยายไปสู่เมทริกซ์นั้นไม่ได้ปิดกั้นโพรงไมโตคอนเดรียอย่างสมบูรณ์และไม่ละเมิดความต่อเนื่องของเมทริกซ์ที่เติมเข้าไป
การวางแนวของคริสเตที่เกี่ยวกับแกนยาวของไมโตคอนเดรียนั้นแตกต่างกันไปตามแต่ละเซลล์ ปฐมนิเทศสามารถตั้งฉาก (ตับไต) คริสตี้; ในกล้ามเนื้อหัวใจมีการจัดเรียงตามยาวของคริสตี้ Crista สามารถแยกหรือสร้างกระบวนการที่มีรูปร่างเป็นนิ้วมืองอและไม่มีทิศทางที่ชัดเจน (รูปที่ 208) ในโปรโตซัวสาหร่ายที่มีเซลล์เดียวในบางเซลล์ของพืชและสัตว์ที่สูงกว่าผลพลอยได้ของเยื่อหุ้มชั้นในจะมีลักษณะคล้ายท่อ (cristae ท่อ)
ไมโตคอนเดรียเมทริกซ์มีโครงสร้างที่เป็นเนื้อเดียวกันซึ่งมีการตรวจพบโมเลกุลดีเอ็นเอในรูปแบบของเส้นใยบาง ๆ ที่เก็บรวบรวมในลูกบอล (ประมาณ 2-3 นาโนเมตร) และไรโบโซมแบบไมโตคอนเดรียในรูปของเม็ดขนาดประมาณ 15-20 นาโนเมตร การสะสมของเกลือแมกนีเซียมและแคลเซียมในเมทริกซ์ก่อให้เกิดเม็ดหนาแน่นขนาดใหญ่ (20-40 นาโนเมตร)
หน้าที่ของไมโตคอนเดรีย
Mitochondria ดำเนินการสังเคราะห์ ATP ซึ่งเกิดขึ้นจากกระบวนการออกซิเดชันของสารตั้งต้นอินทรีย์และ phosphorylation ของ ADP
ขั้นตอนเริ่มต้นของการเกิดออกซิเดชันของคาร์โบไฮเดรตเรียกว่าการเกิดออกซิเดชันแบบไม่ใช้ออกซิเจน (anaerobic oxidation) หรือ glycolysis และเกิดขึ้นใน Hyaloplasm และไม่จำเป็นต้องมีส่วนร่วมของออกซิเจน สารตั้งต้นออกซิเดชั่นระหว่างการผลิตพลังงานแบบไม่ใช้ออกซิเจนคือเฮกโตสและกลูโคส แบคทีเรียบางตัวมีความสามารถในการดึงพลังงานโดยการออกซิไดซ์เพนโตสกรดไขมันหรือกรดอะมิโน
ในกลูโคสปริมาณพลังงานศักย์ที่มีอยู่ในพันธะระหว่างอะตอม C, H และ O มีค่าประมาณ 680 kcal ต่อโมล (เช่น 180 กรัมกลูโคส)
ในเซลล์ที่มีชีวิตพลังงานจำนวนมหาศาลนี้ถูกปล่อยออกมาในรูปแบบของกระบวนการแบบขั้นตอนควบคุมโดยเอนไซม์ออกซิไดซ์จำนวนมากและไม่เกี่ยวข้องกับการเปลี่ยนแปลงของพลังงานพันธะเคมีเข้าสู่ความร้อนเช่นในระหว่างการเผาไหม้ แต่กลายเป็นพันธะแมโครขนาดใหญ่ในโมเลกุล ATP ADP และฟอสเฟต
Trioses เกิดขึ้นจาก glycolysis ส่วนใหญ่กรด pyruvic มีส่วนร่วมในการออกซิเดชั่นต่อไปในไมโทคอนเดรีย ในกรณีนี้การใช้พลังงานของความแตกแยกของพันธะเคมีทั้งหมดเกิดขึ้นซึ่งนำไปสู่การปลดปล่อยของ CO 2 เพื่อการบริโภคของออกซิเจนและการสังเคราะห์ ATP จำนวนมาก กระบวนการเหล่านี้เกี่ยวข้องกับวัฏจักรออกซิเดชันของกรดไตรคาร์บอกซิลิกและห่วงโซ่การหายใจของการถ่ายโอนอิเล็กตรอนโดยที่ phosphorylation ADP และการสังเคราะห์ของเซลล์ "เชื้อเพลิง" และ ATP โมเลกุลเกิดขึ้น (รูปที่ 209)
ในวัฏจักรของกรด tricarboxylic (วงจร Krebs หรือวงจรกรดซิตริก) ไพรูเวตเกิดขึ้นเนื่องจาก glycolysis ครั้งแรกสูญเสียโมเลกุล CO 2 และถูกออกซิไดซ์เป็นอะซิเตท (สารประกอบสองคาร์บอน) รวมกับโคเอนไซม์เอซี รูปแบบหกคาร์บอนซิเตรต (กรดซิตริก) จากนั้นจะมีวัฏจักรของการออกซิเดชั่นของสารประกอบหกคาร์บอนนี้ไปยังออกซาเลตอะซีเตทสี่คาร์บอนอีกครั้งหนึ่งจับกับ acetyl coenzyme A และจากนั้นวัฏจักรจะเกิดซ้ำ ในระหว่างการออกซิเดชั่นนี้จะมีการปล่อย CO 2 โมเลกุลออกไปและอิเล็กตรอนที่ถูกปล่อยออกมาในระหว่างการออกซิเดชั่นจะถูกถ่ายโอนไปยังโมเลกุลโคเอ็นไซม์ตัวรับ (NAD-nicotinamide adenine dinucleotide) ซึ่งเกี่ยวข้องกับพวกมัน ดังนั้นในวงจรของกรดไตรคาร์บอกซิลิกจึงไม่มีการสังเคราะห์เอทีพี แต่มีการเกิดออกซิเดชันของโมเลกุลการถ่ายโอนอิเล็กตรอนไปยังตัวรับและการปล่อย CO 2 เหตุการณ์ทั้งหมดที่อธิบายข้างต้นภายในไมโตคอนเดรียเกิดขึ้นในเมทริกซ์
การออกซิเดชั่นของสารตั้งต้นเริ่มต้นจะนำไปสู่การปลดปล่อย CO 2 และน้ำ แต่พลังงานความร้อนจะไม่ถูกปลดปล่อยออกมาในระหว่างการเผาไหม้ แต่จะเกิดโมเลกุล ATP พวกมันถูกสังเคราะห์โดยโปรตีนกลุ่มอื่นที่ไม่เกี่ยวข้องโดยตรงกับการเกิดออกซิเดชัน ในเยื่อหุ้มไมโตคอนเดรียด้านใน, คอมเพล็กซ์โปรตีนขนาดใหญ่, เอนไซม์, ATP synthetase ตั้งอยู่บนพื้นผิวของเยื่อหุ้มที่มองเข้าไปในเมทริกซ์ ในกล้องจุลทรรศน์อิเล็กตรอนจะมองเห็นได้ในรูปแบบของสิ่งที่เรียกว่า "เห็ด" ร่างกายซึ่งเรียงตามพื้นผิวของเยื่อหุ้มเซลล์อย่างสมบูรณ์มองเข้าไปในเมทริกซ์ ร่างกายมีขาและหัวมีขนาดเส้นผ่านศูนย์กลาง 8-9 นาโนเมตร ดังนั้นเอนไซม์ของทั้งออกซิเดชันโซ่และเอนไซม์สังเคราะห์ ATP จะถูกแปลเป็นภาษาท้องถิ่นในเยื่อหุ้มเซลล์ยล (รูปที่ 201b)
เชนระบบทางเดินหายใจเป็นระบบหลักสำหรับการแปลงพลังงานในไมโตคอนเดรีย ที่นี่การเกิดปฏิกิริยาออกซิเดชั่นแบบต่อเนื่องและการซ่อมแซมองค์ประกอบของห่วงโซ่การหายใจเกิดขึ้นซึ่งเป็นผลมาจากพลังงานที่ถูกปล่อยออกมาในส่วนเล็ก ๆ เนื่องจากพลังงานนี้ที่จุดสามจุดในห่วงโซ่ ATP จึงเกิดจาก ADP และฟอสเฟต ดังนั้นจึงมีการกล่าวว่าออกซิเดชัน (การถ่ายโอนอิเล็กตรอน) มีความเกี่ยวข้องกับฟอสโฟรีเลชั่น (ADP + Fn → ATP, เช่นฟอสโฟรีเลชันออกซิเดชันเกิดขึ้น
พลังงานที่ปล่อยออกมาระหว่างการขนส่งอิเล็กตรอนจะถูกเก็บไว้ในรูปแบบของการไล่ระดับโปรตอนบนเยื่อหุ้มเซลล์ ปรากฎว่าในระหว่างการถ่ายโอนอิเล็กตรอนในเยื่อหุ้มเซลล์ไมโตคอนเดรียแต่ละคอมเพล็กซ์ของระบบทางเดินหายใจสั่งให้พลังงานออกซิเดชันอิสระเคลื่อนที่โปรตอน (ประจุบวก) ผ่านเมมเบรนจากเมทริกซ์ไปยังพื้นที่ intermembrane ซึ่งนำไปสู่การก่อตัวของความแตกต่าง ลบ - จากด้านเมทริกซ์ยล เมื่อถึงความแตกต่างที่อาจเกิดขึ้น (220 mV) โปรตีนคอมเพล็กซ์ของ ATP synthetase เริ่มส่งโปรตอนกลับไปที่เมทริกซ์ในขณะที่เปลี่ยนพลังงานรูปแบบหนึ่งไปเป็นอีกรูปแบบหนึ่ง: สร้าง ATP จาก ADP และอนินทรีย์ฟอสเฟต นี่คือการรวมกันของกระบวนการออกซิเดชั่นกับสังเคราะห์กับฟอสโฟรีเลชั่นของ ADP ในขณะที่สารตั้งต้นออกซิไดซ์ในขณะที่โปรตอนถูกสูบผ่านเยื่อหุ้มไมโตคอนเดรียลชั้นในการสังเคราะห์ ATP ที่เกี่ยวข้องกับสิ่งนี้เกิดขึ้นคือ เกิดออกซิเดชัน phosphorylation
กระบวนการทั้งสองนี้สามารถแยกออกจากกัน ในกรณีนี้การถ่ายโอนอิเล็กตรอนจะดำเนินต่อไปเช่นเดียวกับการออกซิเดชั่นของสารตั้งต้น แต่การสังเคราะห์ ATP จะไม่เกิดขึ้น ในกรณีนี้พลังงานที่ปล่อยออกมาระหว่างออกซิเดชั่นจะถูกแปลงเป็นพลังงานความร้อน
phosphorylation ออกซิเดชันในแบคทีเรีย.
ในเซลล์โปรคาริโอตที่มีความสามารถในการออกซิเดชั่นฟอสโฟรีเลชั่นองค์ประกอบของวัฏจักรกรด tricarboxylic จะถูกแปลโดยตรงในไซโตพลาสซึมและห่วงโซ่การหายใจและเอนไซม์ฟอสโฟรีเลชั่นเกี่ยวข้องกับเยื่อหุ้มเซลล์ มันควรจะตั้งข้อสังเกตว่าแบคทีเรีย mesosomes ไม่เพียง แต่เกี่ยวข้องกับกระบวนการหายใจแอโรบิก แต่ในบางชนิดมีส่วนร่วมในการแบ่งเซลล์ในกระบวนการกระจายดีเอ็นเอในเซลล์ใหม่ในการก่อตัวของผนังเซลล์ ฯลฯ
บนพลาสมาเมมเบรนใน mesosomes ของแบคทีเรียบางชนิดกระบวนการเกิดปฏิกิริยาผันแปรระหว่างการเกิดออกซิเดชันและการสังเคราะห์ ATP อนุภาคทรงกลมคล้ายกับที่พบในไมโตคอนเดรียของเซลล์ยูคาริโอตถูกพบในกล้องจุลทรรศน์อิเล็กตรอนในส่วนของเยื่อหุ้มพลาสม่าของแบคทีเรีย ดังนั้นในเซลล์แบคทีเรียที่มีความสามารถในการออกซิเดทีฟฟอสโฟรีเลชั่นเมมเบรนในพลาสมามีบทบาทคล้ายกับเยื่อหุ้มยลภายในของเซลล์ยูคาริโอต
การเพิ่มขึ้นของจำนวนไมโตคอนเดรีย
Mitochondria สามารถเพิ่มจำนวนของพวกเขาโดยเฉพาะอย่างยิ่งกับการแบ่งเซลล์หรือเพิ่มภาระการทำงานของเซลล์ มีการต่ออายุของไมโตคอนเดรียอย่างต่อเนื่อง ตัวอย่างเช่นในตับช่วงชีวิตเฉลี่ยของไมโตคอนเดรียประมาณ 10 วัน
การเพิ่มขึ้นของจำนวนไมโตคอนเดรียเกิดขึ้นผ่านการเติบโตและการแบ่งไมโตคอนเดรียก่อนหน้า ข้อสันนิษฐานนี้ถูกแสดงออกมาเป็นครั้งแรกโดยอัลท์แมน (1893) ซึ่งอธิบายถึงไมโตคอนเดรียภายใต้คำว่า "ไบโอกลาสต์" มันเป็นไปได้ที่จะสังเกตในการแบ่งวิฟการแยกส่วนของไมโทคอนเดรียยาวเป็นแบบสั้นโดยการหดตัวซึ่งคล้ายกับวิธีไบนารีของการแบ่งแบคทีเรีย
การเพิ่มจำนวนของไมโตคอนเดรียอย่างแท้จริงโดยการหารนั้นเกิดขึ้นเมื่อศึกษาพฤติกรรมของไมโตคอนเดรียในเซลล์ที่มีชีวิตของการเพาะเลี้ยงเนื้อเยื่อ ในช่วงวัฏจักรของเซลล์ไมโตคอนเดรียจะเติบโตเป็นหลายไมครอนแล้วแยกส่วนออกเป็นส่วนย่อย ๆ
Mitochondria สามารถรวมเข้าด้วยกันและทวีคูณตามหลักการ: Mitochondria จาก Mitochondria
Autoproduction ของไมโตคอนเดรีย
organelles สองเมมเบรนมีระบบการทำสำเนาอัตโนมัติที่สมบูรณ์ ในไมโทคอนเดรียและ plastids มี DNA ที่ข้อมูลการถ่ายโอนและไรโบโซม RNAs และไรโบโซมถูกสังเคราะห์ซึ่งดำเนินการสังเคราะห์โปรตีนไมโตคอนเดรียและพลาสมิด อย่างไรก็ตามระบบเหล่านี้แม้ว่าจะเป็นอิสระ แต่ก็มีข้อ จำกัด ในความสามารถของพวกเขา
ดีเอ็นเอในไมโทคอนเดรียเป็นโมเลกุลแบบไซโคลที่ไม่มีฮิสโตสและคล้ายกับโครโมโซมจากแบคทีเรีย ขนาดของมันคือประมาณ 7 ไมครอน, 16-19,000 คู่นิวคลีโอไทด์ของดีเอ็นเอเข้าสู่โมเลกุลวงจรหนึ่งของสัตว์ไมโทคอนเดรีย ในมนุษย์ DNA ไมโตคอนเดรียมี 16.5 พัน n.p. ซึ่งถอดรหัสได้อย่างสมบูรณ์ พบว่า DNA ไมโตคอนดรัลของวัตถุต่าง ๆ นั้นเป็นเนื้อเดียวกันมากความแตกต่างของพวกมันอยู่ในขนาดของอินตรอนและไซต์ที่ไม่ถูกถ่ายเท่านั้น ไมโตคอนเดรียดีเอ็นเอทั้งหมดนั้นมีหลาย ๆ กลุ่มรวมกันเป็นกลุ่ม ดังนั้นหนึ่งในไมโตคอนเดรียของตับหนูสามารถมีได้ตั้งแต่ 1 ถึง 50 โมเลกุลดีเอ็นเอ จำนวน mitochondrial DNA ทั้งหมดต่อเซลล์นั้นประมาณหนึ่งเปอร์เซ็นต์ การสังเคราะห์ดีเอ็นเอไมโตคอนเดรียไม่เกี่ยวข้องกับการสังเคราะห์ดีเอ็นเอในนิวเคลียส
เช่นเดียวกับในแบคทีเรีย mitochondral DNA จะถูกรวบรวมในโซนแยกต่างหาก - นิวรอยด์ขนาดของมันมีขนาดเส้นผ่าศูนย์กลางประมาณ 0.4 ไมครอน ในไมโทคอนเดรียที่ยาวนานสามารถมีนิวรอยด์ได้ตั้งแต่ 1 ถึง 10 เมื่อแบ่ง mitochondria ที่มีความยาวส่วนที่มี nucleoid จะถูกแยกออกจากมัน (คล้ายกับการแบ่งไบนารีของแบคทีเรีย) ปริมาณของ DNA ในนิวครอยด์ไมโตคอนเดรียแต่ละตัวสามารถผันผวนได้ 10 ครั้งขึ้นอยู่กับชนิดของเซลล์
ในบางวัฒนธรรมในเซลล์จาก 6 ถึง 60% ของไมโทคอนเดรียไม่มีนิวเคลียสซึ่งสามารถอธิบายได้ด้วยความจริงที่ว่าการแบ่งตัวของออร์แกเนลล์เหล่านี้มีความเกี่ยวข้องกับการกระจายตัวมากกว่าการกระจายตัวของนิวรอยด์
ดังที่ได้กล่าวไปแล้วไมโตคอนเดรียสามารถแบ่งและรวมเข้าด้วยกัน เมื่อไมโตคอนเดรียผสานเข้าด้วยกันการแลกเปลี่ยนของชิ้นส่วนภายในอาจเกิดขึ้นได้
สิ่งสำคัญคือต้องเน้นว่า rRNA และไรโบโซมของไมโทคอนเดรียและไซโตพลาสซึมแตกต่างกันอย่างมาก หากไรโบโซม 80s ถูกพบในไซโตพลาสซึมแล้วไรโบโซมยลของเซลล์พืชจะอยู่ในไรโบโซม 70s (ประกอบด้วย 30s และ 50s ย่อย, มี 16 และ 23s RNAs ของเซลล์โปรคาริโอตตี้) และไรโบโซมขนาดเล็ก
Mitochondrial ribosomal RNAs ถูกสังเคราะห์บนยลไมโตคอนเดรีย ใน mitoplasm บนไรโบโซมเป็นการสังเคราะห์โปรตีน มันหยุดตรงกันข้ามกับการสังเคราะห์บนไรโบโซมของไซโตพลาสซึมภายใต้การกระทำของยาปฏิชีวนะคลอแรมเฟนิคอลซึ่งยับยั้งการสังเคราะห์โปรตีนในแบคทีเรีย
ในจีโนม mitochondrial, 22 ขนส่ง RNAs จะถูกสังเคราะห์ รหัส triplet ของระบบสังเคราะห์ยลนั้นแตกต่างจากที่ใช้ในการทำไฮยาโลพลาสม่า แม้จะมีส่วนประกอบทั้งหมดที่ดูเหมือนจะจำเป็นสำหรับการสังเคราะห์โปรตีน แต่โมเลกุลดีเอ็นเอไมโตคอนเดรียขนาดเล็กไม่สามารถเข้ารหัสโปรตีนไมโตคอนเดรียทั้งหมดได้เพียงส่วนเล็ก ๆ เท่านั้น ดังนั้นดีเอ็นเอในขนาด 15,000 n.p. สามารถเข้ารหัสโปรตีนที่มีน้ำหนักโมเลกุลรวมประมาณ 6x10 5 ในเวลาเดียวกันนั้นน้ำหนักโมเลกุลทั้งหมดของโปรตีนของอนุภาคของไมโทคอนเดรียทางเดินหายใจทั้งหมดมีค่าประมาณ 2x10 6
เมื่อพิจารณาว่านอกเหนือไปจากโปรตีนออกซิเดทีฟฟอสโฟรีเลชั่นแล้วไมโทคอนเดรียยังรวมถึงเอนไซม์วัฏจักรกรดไตรคาร์บอกซิลิกเอนไซม์การสังเคราะห์ DNA และ RNA เอนไซม์กระตุ้นการทำงานของกรดอะมิโนและโปรตีนอื่น ๆ โมเลกุลของไมโทคอนเดรียสั้นนั้นขาดชัดเจน การถอดรหัสลำดับนิวคลีโอไทด์ของ DNA ยลยลของมนุษย์แสดงให้เห็นว่ามันเข้ารหัสเพียง 2 ribosomal RNAs, 22 ถ่ายโอน RNAs และทั้งหมด 13 โซ่โพลีเปปไทด์ที่แตกต่างกัน
ตอนนี้ได้รับการพิสูจน์แล้วว่าโปรตีนส่วนใหญ่ของไมโตคอนเดรียอยู่ภายใต้การควบคุมทางพันธุกรรมจากนิวเคลียสของเซลล์และถูกสังเคราะห์นอกไมโตคอนเดรีย โปรตีนส่วนใหญ่จะถูกสังเคราะห์บนไรโบโซมในไซโตทอล โปรตีนเหล่านี้มีลำดับสัญญาณพิเศษที่ได้รับการยอมรับจากตัวรับบนเยื่อหุ้มเซลล์ยล โปรตีนเหล่านี้สามารถแทรกเข้าไปในพวกมัน (ดูการเปรียบเทียบกับเมมเบรนรอกซ์ซิโซม) จากนั้นย้ายไปที่เยื่อหุ้มชั้นใน การถ่ายโอนนี้เกิดขึ้นที่จุดสัมผัสของเยื่อหุ้มด้านนอกและด้านในซึ่งมีการทำเครื่องหมายการขนส่งดังกล่าว ไขมันส่วนใหญ่จะถูกสังเคราะห์ในไซโตพลาสซึม
ทั้งหมดนี้บ่งบอกถึงต้นกำเนิดของเอนโดซิมไบโอติกของไมโทคอนเดรียซึ่งไมโทคอนเดรียเป็นสิ่งมีชีวิตเช่นแบคทีเรียที่อยู่ใน symbiosis กับเซลล์ยูคาริโอต
chondrite
จำนวนทั้งสิ้นของไมโตคอนเดรียทั้งหมดในเซลล์เดียวเรียกว่า chondrioma มันอาจแตกต่างกันไปขึ้นอยู่กับประเภทของเซลล์ ในเซลล์จำนวนมาก chondriomas ประกอบด้วยไมโทคอนเดรียจำนวนมากกระจายอยู่ทั่วทั้งไซโตพลาสซึมหรือแปลเป็นกลุ่มในไซต์ที่มีการใช้จ่าย ATP อย่างเข้มข้น ในทั้งสองกรณีนี้ไมโตคอนเดรียทำงานเพียงอย่างเดียวทำงานร่วมกันอาจมีสัญญาณบางส่วนจากไซโตพลาสซึม มีประเภทของ chondrioma ที่แตกต่างกันอย่างสิ้นเชิงเมื่อแทนที่จะเป็นไมโตคอนเดรียกระจัดกระจายเดี่ยวไมโตคอนเดรียยักษ์ตัวหนึ่งจะอยู่ในเซลล์
ไมโตคอนเดรียดังกล่าวพบในสาหร่ายสีเขียวที่มีเซลล์เดียว (ตัวอย่างเช่น Chlorella) พวกมันก่อตัวเป็น mitochondrial network ที่ซับซ้อนหรือ reticulum mitochondrial (Reticulum miyochondriale) ตามทฤษฎี chemo-osmotic ความหมายทางชีวภาพของการปรากฏตัวของโครงสร้าง mitochondrial ยักษ์ที่แยกจากกันโดยเยื่อหุ้มภายนอกและภายในคือการสังเคราะห์ ATP สามารถเกิดขึ้นได้ทุกจุดบนพื้นผิวของเยื่อหุ้มชั้นในของ mitochondria ซึ่งจะไปที่ใดก็ได้ในไซโตพลาสซึม ไม่จำเป็นต้องมีสิ่งนี้
ในกรณีของไมโทคอนเดรียแขนงยักษ์ ณ จุดใด ๆ มันก็มีศักยภาพเพียงพอที่จะเริ่มการสังเคราะห์เอทีพีสามารถสะสมบนเยื่อหุ้มชั้นใน จากตำแหน่งเหล่านี้ reticulum mitochondrial คืออย่างที่มันเป็นตัวนำไฟฟ้าสายเคเบิลที่เชื่อมต่อจุดที่ห่างไกลของระบบดังกล่าว mitochondrial reticulum นั้นมีประโยชน์มากไม่เพียง แต่สำหรับเซลล์เคลื่อนที่ขนาดเล็กเช่นคลอเรลล่า แต่ยังสำหรับหน่วยโครงสร้างขนาดใหญ่เช่น myofibrils ในกล้ามเนื้อโครงร่าง
มันเป็นที่รู้จักกันว่ากล้ามเนื้อโครงร่างประกอบด้วยมวลของเส้นใยกล้ามเนื้อ symplasts ที่มีนิวเคลียสจำนวนมาก ความยาวของเส้นใยกล้ามเนื้อเช่นนี้ถึง 40 ไมครอนมีความหนา 0.1 ไมครอน - นี่คือโครงสร้างขนาดมหึมาที่มี myofibrils จำนวนมากซึ่งทั้งหมดทำสัญญาพร้อมกันพร้อมกัน สำหรับการหดตัว ATP จำนวนมากจะถูกส่งไปยังแต่ละหน่วยหดตัว myofibril ซึ่ง mitochondria จัดเตรียมไว้ที่ระดับของ z-disk ในส่วน ultrathin ultrathin ยาวของกล้ามเนื้อโครงกระดูกกล้องจุลทรรศน์อิเล็กตรอนจะแสดงส่วนเล็ก ๆ หลาย ๆ อันของไมโทคอนเดรียที่อยู่ในบริเวณใกล้เคียงของ sarcomeres ไมโทคอนเดรียกล้ามเนื้อไม่ใช่ลูกบอลหรือแท่งเล็ก ๆ แต่มีโครงสร้างแบบแมงซึ่งเป็นกระบวนการที่แตกกิ่งก้านสาขาและขยายออกไปเป็นระยะทางไกลบางครั้งมีเส้นผ่านศูนย์กลางทั้งหมดของเส้นใยกล้ามเนื้อ
ในเวลาเดียวกันนั้นการแยกไมโตคอนเดรียรอบ myofibril แต่ละอันในเส้นใยกล้ามเนื้อทำให้ ATP นั้นจำเป็นสำหรับการหดตัวของกล้ามเนื้อ ดังนั้นในระนาบของ z-disk mitochondria จึงเป็นตัวแทนของ reticulum แบบไมโตคอนเดรียทั่วไป mitochondrial reticulum ดังกล่าวซ้ำสองครั้งสำหรับแต่ละ sarcomere และเส้นใยกล้ามเนื้อทั้งหมดมีพัน "ชั้น" ขวางอยู่ของชั้นของ mitochondrial reticulum ระหว่าง "พื้น" ตาม myofibrils เป็นเส้นใยไมโตคอนเดรียเชื่อมต่อเลเยอร์ยลเหล่านี้ ดังนั้นภาพสามมิติของเรติเคิลไมโทคอนเดรียซึ่งผ่านเส้นใยกล้ามเนื้อในปริมาณทั้งหมดจึงถูกสร้างขึ้น
มันเป็นที่ยอมรับต่อไปว่าระหว่างกิ่งก้านของไมโทคอนเดรียลและไมโทคอนเดรียยาวตามแนวยาวมีเส้นใย intermitochondrial สารประกอบพิเศษหรือการติดต่อ (MMC) พวกมันเกิดขึ้นจากเยื่อหุ้มไมโตคอนเดรียที่อยู่ติดกันอย่างแน่นหนาบริเวณที่สัมผัสกับไมโตคอนเดรียพื้นที่ระหว่างเมมเบรนและเมมเบรนในโซนนี้มีความหนาแน่นของอิเล็กตรอนเพิ่มขึ้น ด้วยการก่อตัวพิเศษเหล่านี้สหภาพการทำงานของไมโทคอนเดรียและเรติเคิลโทมิกซึ่งอยู่ใกล้เคียงจะเกิดขึ้นในระบบพลังงานสหกรณ์เพียงระบบเดียว myofibrils ทั้งหมดในเส้นใยกล้ามเนื้อจะลดลงพร้อมกันตลอดความยาวดังนั้นอุปทานของ ATP ในส่วนใด ๆ ของเครื่องที่ซับซ้อนนี้ก็ควรจะเกิดขึ้นพร้อมกันและสิ่งนี้จะเกิดขึ้นได้ถ้ามีตัวนำ mitochondria แยกกันจำนวนมากเท่านั้น ใช้ที่อยู่ติดต่อ
ความจริงที่ว่าการติดต่อระหว่างกัน (MMK) มีส่วนร่วมในสมาคมพลังงานของไมโทคอนเดรียซึ่งกันและกันเป็นไปได้ใน cardiomyocytes, เซลล์กล้ามเนื้อหัวใจ
Chondriomas ของเซลล์กล้ามเนื้อหัวใจไม่ได้สร้างโครงสร้างการแตกแขนง แต่จะถูกแทนด้วยไมโตคอนเดรียขนาดเล็กจำนวนมากที่ตั้งอยู่โดยไม่มีคำสั่งพิเศษระหว่าง myofibrils อย่างไรก็ตามไมโตคอนเดรียทั้งหมดที่อยู่ใกล้เคียงเข้าด้วยกันด้วยความช่วยเหลือจากไมโทคอนเดรียชนิดเดียวกับกล้ามเนื้อโครงร่างมีเพียงจำนวนมาก: โดยเฉลี่ยมี 2-3 เอ็มซีต่อ Mitochondria ซึ่งผูกไมโทคอนเดรียเข้าด้วยกัน ห่วงโซ่ (Streptio mitochondriale) เป็น mitochondria แยกต่างหาก
มันปรากฏว่าการติดต่อระหว่างกัน (MMC) ในฐานะโครงสร้างบังคับของเซลล์หัวใจถูกพบใน cardiomyocytes ของโพรงและ atria ของสัตว์มีกระดูกสันหลังทั้งหมด: สัตว์เลี้ยงลูกด้วยนมนกสัตว์เลื้อยคลานสัตว์ครึ่งบกครึ่งน้ำและปลากระดูก ยิ่งกว่านั้นพบ MMCs (แต่ในจำนวนที่น้อยกว่า) ในเซลล์หัวใจของแมลงและหอยบางชนิด
ปริมาณ MMC ใน cardiomyocytes จะแตกต่างกันไปขึ้นอยู่กับภาระการทำงานของหัวใจ จำนวน MMC เพิ่มขึ้นตามการออกกำลังกายที่เพิ่มขึ้นของสัตว์และในทางกลับกันเมื่อปริมาณกล้ามเนื้อหัวใจลดลงจำนวน MMC ก็ลดลงอย่างรวดเร็ว