Tā kā tiek sauktas mitohondriju iekšējās struktūras. Mitohondriju un plastižu struktūra un funkcijas
Mitohondriji
Dzīvnieku audu šūnās mitohondriji tika atrasti 1882. gadā, bet augos - tikai 1904. gadā (ūdensrožu skudrās). Pēc frakcijas atdalīšanas un attīrīšanas ar frakcionētu centrifugēšanu bija iespējams noteikt bioloģiskās funkcijas. Tie satur 70% olbaltumvielu un apmēram 30% lipīdu, nelielu daudzumu RNS un DNS, vitamīnus A, B 6, B 12, K, E, folijskābes un pantotēnskābes, riboflavīnu, dažādus fermentus. Mitohondrijiem ir dubultā membrāna, ārējā izolē organellu no citoplazmas, un iekšējā veido krista izaugumus. Visa telpa starp membrānām ir piepildīta ar matricu (13. att.).
Mitohondriju galvenā funkcija ir dalība šūnu elpošanā. Mitohondriju loma elpošanā tika noteikta 1950.-1951. Krebsa cikla kompleksa enzīmu sistēma ir koncentrēta uz ārējām membrānām. Elpošanas substrātu oksidācijas laikā tiek atbrīvota enerģija, kas nekavējoties tiek uzkrāta izveidotajās ADP molekulās, galvenokārt ATP, oksidējošās fosforilēšanās laikā, kas notiek kraukšķī. Makroerģiskos savienojumos uzkrātā enerģija nākotnē tiek izmantota, lai apmierinātu visas šūnas vajadzības.
Mitohondriju veidošanās šūnā notiek nepārtraukti no mikroorganismiem, biežāk to rašanās ir saistīta ar šūnas membrānu struktūru diferenciāciju. Tos var atjaunot šūnā, tos sadalot un ieliekot. Mitohondriji nav izturīgi, viņu dzīves ilgums ir 5-10 dienas.
Mitohondriji ir “spēka” šūnu stacijas. Tie koncentrē enerģiju, kas tiek glabāta enerģijas "akumulatoros" - ATP molekulās un netiek izkliedēta šūnā. Mitohondriju struktūras pārkāpums noved pie elpošanas procesa traucējumiem un rezultātā ķermeņa patoloģijām.
Golgi aparāts.Golgi aparāts (sinonīms - dicitosomas) ir 3–12 kaudze, saplacināta, slēgta, ko ieskauj dubultā membrānas diski, kurus sauc par tvertnēm, no kuru malām ir nesaspringti daudzi burbuļi (300–500). Tvertņu platums ir 6-90 A, membrānu biezums ir 60-70 A.
Golgi aparāts ir polisaharīdu, jo īpaši celulozes, sintēzes, uzkrāšanās un izolācijas centrs, un tas ir iesaistīts olbaltumvielu izplatīšanā un intracelulārā transportēšanā, kā arī vakuolu un lizosomu veidošanā. Augu šūnā bija iespējams izsekot Golgi aparāta līdzdalībai vidējās plāksnes izskatā un šūnas pektocelulozes membrānas augšanā.
Golgi aparāts ir visattīstītākais šūnu aktīvās dzīves periodā. Tā kā tas noveco, tas pakāpeniski atrofējas un pēc tam pazūd.
Lizosomas.Lizosomas - diezgan mazi (apmēram 0,5 mikronu diametrā) noapaļoti korpusi. Tie ir pārklāti ar olbaltumvielu lipoīdu membrānu. Lizosomu saturs ir daudz hidrolītisku enzīmu, kas veic olbaltumvielu makromolekulu, nukleīnskābju, polisaharīdu intracelulāras šķelšanas (sabrukšanas) funkciju. Viņu galvenā funkcija ir šūnu protoplastu atsevišķu sekciju (autofagija - pašnodarbināšanās) gremošana. Šis process norit fagocitozes vai pinocitozes dēļ. Šim procesam ir divējāda loma. Pirmkārt, tā ir aizsargājoša, jo ar īslaicīgu rezerves produktu trūkumu šūna atbalsta dzīvību konstitucionālo olbaltumvielu un citu vielu dēļ, un, otrkārt, atbrīvojas no liekiem vai nolietotiem organelliem (plastidiem, mitohondrijiem utt.). Lizosomu membrāna novērš enzīmu izdalīšanos citoplazmā. pretējā gadījumā šie fermenti to pilnībā sagremotu.
Atmirušajā šūnā tiek iznīcinātas lizosomas, šūnā atrodas fermenti un viss tā saturs tiek sagremots. Atliek tikai petrocelulozes apvalks.
Lizosomas ir Golgi aparāta darbības produkti, no tā atdalīti pūslīši, kuros šis organoīds uzkrāja gremošanas fermentus.
Sferosomas - noapaļoti olbaltumvielu-lipoīdu ķermeņi 0,3–0,4 mikroni. Visticamāk, tie ir Golgi aparāta vai endoplazmas retikuluma atvasinājumi. Pēc formas un izmēra atgādina lizosomas. Tā kā sferosomas satur skābo fosfatāzi, tās, iespējams, ir saistītas ar lizosomām. Daži autori uzskata, ka sferosomas un lizosomas ir līdzvērtīgas viena otrai, bet, visticamāk, tikai pēc izcelsmes un formas. Pastāv pieņēmums par viņu līdzdalību tauku sintēzē (A. Frey-Wissling).
Ribosomas - ļoti mazi organoīdi, to diametrs ir apmēram 250 A. Tie ir gandrīz sfēriski. Daži no tiem ir piestiprināti pie endoplazmatiskā retikulārā auduma ārējām membrānām, daži no tiem ir brīvā stāvoklī citoplazmā. Šūna var saturēt līdz 5 miljoniem ribosomu. Ribosomas ir atrodamas hloroplastos un mitohondrijos, kur tie sintezē daļu no olbaltumvielām, kurās šie organoīdi ir veidoti, un fermentus, kas tajās darbojas.
Galvenā funkcija ir specifisku olbaltumvielu sintēze saskaņā ar informāciju no kodola. To sastāvs: olbaltumviela un ribosomāla ribonukleīnskābe (RNS) vienādās proporcijās. Viņu struktūra ir maza un liela subvienība, kas veidojas no ribonukleotīda.
Mikrotubulas.Mikrotubulas - savdabīgi endoplazmatiskā retikuluma atvasinājumi. Atrasts daudzās šūnās. Viņu vārds pats par sevi runā par viņu formu - vienu vai diviem, kas atrodas paralēli, kanāliņos ar dobumu iekšpusē. Ārējais diametrs ir 250A. Mikrotubulu sienas ir veidotas no olbaltumvielu molekulām. Šūnu dalīšanas laikā no mikrotubuliem veidojas vārpstas pavedieni.
Kodols
Kodolu augu šūnā atklāja R. Brauns 1831. gadā. Tas atrodas šūnas centrā vai netālu no šūnas membrānas, bet no visām pusēm to ieskauj citoplazma. Vairumā gadījumu šūnā ir viens kodols, vairāku kodolu atrodas dažu aļģu, kā arī sēnīšu šūnās. Bezšūnu zaļajām aļģēm ir simtiem kodolu. Nesavienotu laktēru daudzkodolu šūnas. Baktēriju un zili zaļo aļģu šūnās nav kodolu.
Kodola forma visbiežāk ir tuvu bumbiņas vai elipses formai. Atkarīgs no šūnas formas, vecuma un funkcijas. Meristematiskajā šūnā kodols ir liels, apaļas formas un aizņem 3/4 no šūnas tilpuma. Epidermas parenhīmas šūnās ar lielu centrālo vakuolu kodolam ir lencveida forma un tas kopā ar citoplazmu tiek pārvietots uz šūnas perifēriju. Tas liecina par specializētu, bet jau novecojošu šūnu. Šūnā, kurā nav kodola, var dzīvot tikai īsu laiku. Sietu caurulīšu šūnas, kas nesatur kodolu, ir dzīvas šūnas, taču tās nedzīvo ilgi. Visos citos gadījumos šūnas bez kodoliem ir mirušas.
Kodolam ir dubultā čaula caur porām, kurās ir saturs
kodoli (nukleoplazma) var sazināties ar citoplazmas saturu. Kodola membrānas ir aprīkotas ar ribosomām un sazinās ar šūnas endoplazmatiskā retikulāra membrānām. Nukleoplazmā atrodas viena vai divas nukleoli un hromosomas. Kodolplazma ir Zola koloidālā sistēma, kas pēc konsistences atgādina sabiezinātu želatīnu. Pēc krievu bioķīmiķu (Zbarsky I.B. et al.) Domām, kodolā ir četras olbaltumvielu frakcijas: vienkāršie proteīni - globulīni 20%, dezoksiribonukleoproteīni - 70%, skābes olbaltumvielas - 6% un atlikušie proteīni 4%. Tie ir lokalizēti šādās kodola struktūrās: DNS proteīdi (sārmainās olbaltumvielas) hromosomās, RNS proteīdi (skābie proteīni) nukleolos, daļēji hromosomās (MNS sintēzes laikā) un kodolenerģijas membrānā. Globulīni veido nukleoplazmas pamatu. Atlikušie olbaltumvielas (raksturs nav noteikts) veido kodola membrānu.
Lielākā daļa galveno olbaltumvielu ir kompleksi sārmaini proteīni, dezoksiribonukleoproteīni, kuru pamatā ir DNS.
DNS molekula.DNS molekula - polinukleotīds un sastāv no nukleotīdiem. Nukleotīds satur trīs komponentus: cukura molekulu (dezoksiribozi), slāpekļa molekulu un fosforskābes molekulu. Dezoksiriboze ir saistīta ar slāpekļa bāzes glikozīdisko bāzi, un ar fosforskābi - ētera saiti. DNS dažādās kombinācijās ir tikai 4 nukleotīdu varianti, kas viens no otra atšķiras slāpekļa bāzēs. Divas no tām (adenīns un guanīns) pieder purīniem slāpekļa savienojumiem, bet citozīns un timīns - pirimidīnam. DNS molekulas neatrodas vienā plaknē, bet sastāv no divām spirālveida šķipsnām, t.i. divas paralēlas ķēdes, kas savītas viena otrai apkārt, veido vienu DNS molekulu. Tās ir savstarpēji saistītas ar ūdeņraža saiti starp slāpekļa bāzēm, un vienas ķēdes purīna bāzes piestiprina otras puses pirimidīna bāzes (14. att.). DNS molekulas struktūru un ķīmiju atklāja angļu (Kriks) un Amerikas (Vatsons) zinātnieki, un to publiskoja 1953. gadā. Šis punkts tiek uzskatīts par molekulārās ģenētikas attīstības sākumu. DNS molekulmasa ir 4–8 miljoni.Nukleotīdu (dažādu variantu) skaits ir līdz 100 tūkstošiem. DNS molekula ir ļoti stabila, tās stabilitāti nodrošina tas, ka visā tajā ir vienāds biezums - 20A (8A - pirimidīna bāzes platums + 12A - purīna bāzes platums). Ja ķermenī tiek ievadīts radioaktīvais fosfors, marķējums tiks atklāts visiem fosforu saturošiem savienojumiem, izņemot DNS (Levy, Sickewitz).
DNS molekulas ir iedzimtības nesēji, jo to struktūra kodē informāciju par īpašu olbaltumvielu sintēzi, kas nosaka ķermeņa īpašības. Izmaiņas var notikt mutagēnu faktoru ietekmē (radioaktīvais starojums, spēcīgi, ķīmiski līdzekļi - alkaloīdi, spirti utt.).
RNS molekula.Ribonukleīnskābes molekulas (RNS) ievērojami mazāk DNS molekulu. Tās ir vienas nukleotīdu ķēdes. Pastāv trīs RNS veidi: ribosomāli, garākie, veidojot daudzas cilpas, informācija (matrica) un īsākie. Ribosomāla RNS ir lokalizēta endoplazmatiskā retikuluma ribosomās un veido 85% no kopējās šūnas RNS.
Informatīvā RNS savā struktūrā atgādina āboliņa lapu. Tās daudzums ir 5% no kopējās RNS šūnā. Tas tiek sintezēts nukleolos. Tās montāža notiek hromosomās starpfāzu periodā. Tās galvenā funkcija ir informācijas pārnešana no DNS uz ribosomām, kur notiek olbaltumvielu sintēze.
Transporta RNS, kā tas tagad ir noteikts, ir vesela savienojumu grupa, kas saistīta ar struktūru un bioloģisko funkciju. Katrā dzīvā šūnā, pēc aptuveniem aprēķiniem, ir 40-50 individuālo transporta RNS, un to kopējais skaits dabā, ņemot vērā sugu atšķirības, ir milzīgs. (Akad. V. Engelhards). Tos sauc par transportu, jo to molekulas nodarbojas ar olbaltumvielu sintēzes intracelulārā procesa transporta uzturēšanu. Apvienojumā ar brīvajām aminoskābēm tās piegādā proteīna ķēdē esošajās ribosomās. Šīs ir mazākās RNS molekulas, kas sastāv vidēji no 80 nukleotīdiem. Lokalizēts citoplazmas matricā un veido apmēram 10% no šūnu RNS
RNS satur četras slāpekļa bāzes, taču atšķirībā no DNS RNS timīna vietā ir uracils.
Hromosomu struktūra.Hromosomas pirmo reizi 19. gadsimta beigās atklāja citoloģijas klasiķi Flemings un Strasburgers (1882, 1884) un krievu šūnu pētnieks I.D. Čistjakovs tos atklāja 1874. gadā.
Hromosomu galvenais strukturālais elements ir kodols. Viņiem ir atšķirīga forma. Tie ir vai nu taisni, vai izliekti spieķi, ovāli korpusi, bumbiņas, kuru izmēri atšķiras.
Atkarībā no centromēra atrašanās vietas tiek izdalītas tiešās, vienādās un nevienādās hromosomas. Hromosomu iekšējā struktūra ir parādīta att. 15, 16. Jāatzīmē, ka dezoksiribonukleoproteīns ir hromosomas monomērs.
Dezoksiribonukleoproteīnu hromosomā 90–92%, no kuriem 45% ir DNS un 55% ir olbaltumvielas (histons). RNS (informatīvs) nelielā daudzumā ir pārstāvēts arī hromosomā.
Hromosomai ir arī izteikta šķērseniska struktūra - sabiezinātu zonu klātbūtne - diski, kas bija atpakaļ 1909. gadā. sauca par gēniem. Šo terminu ierosināja dāņu zinātnieks Johansens. 1911. gadā amerikāņu zinātnieks Morgans pierādīja, ka gēni ir galvenās iedzimtās vienības un tie hromosomās tiek sadalīti lineārā secībā, un tāpēc hromosomā ir kvalitatīvi atšķirīgas sadaļas. 1934. gadā amerikāņu zinātnieks Pinters pierādīja hromosomu morfoloģiskās struktūras pārtraukumu un disku klātbūtni hromosomās, un diski ir vietas, kur uzkrājas DNS. Tas kalpoja par sākumu hromosomu karšu veidošanai, kas norādīja gēna atrašanās vietu (lokusu), kas nosaka vienu vai otru organisma iezīmi. Gēns ir DNS dubultās spirāles reģions, kas satur informāciju par atsevišķa proteīna struktūru. Šī ir DNS molekulas sadaļa, kas nosaka vienas olbaltumvielu molekulas sintēzi. DNS tieši nepiedalās olbaltumvielu sintēzē. Tas satur un glabā tikai informāciju par olbaltumvielu struktūru.
DNS struktūra, kas sastāv no vairākiem tūkstošiem secīgi izvietotu 4 nukleotīdu, ir iedzimtības kods.
Iedzimtības kods. Olbaltumvielu sintēze.Pirmo ziņojumu par DNS kodu amerikāņu bioķīmiķis Nirenbergs sniedza 1961. gadā Maskavā Starptautiskajā bioķīmiskajā kongresā. DNS koda būtība ir šāda. Katra aminoskābe atbilst triju blakus esošo nukleotīdu (tripletu) DNS ķēdes daļai. Tātad, piemēram, vietne, kas sastāv no TT-T (trīs timīnu saturošu nukleotīdu trīskāršs), atbilst aminoskābei lizīnam, tripletam A (adenīns) - C (citozīns) - A (adenīnam) - cisteīnam utt. Pieņemsim, ka gēnu attēlo nukleotīdu virkne, kas sakārtota šādā secībā: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G. Sadalot šo sēriju trīskāršos, mēs nekavējoties atšifrēsim, kuras aminoskābes un kādā secībā atradīsies sintezētajā olbaltumvielā.
Iespējamo 4 pieejamo trīs nukleotīdu trīs kombināciju skaits ir 4 × 64. Balstoties uz šīm attiecībām, dažādu trīskāršojošo skaita pārsniegums būs pietiekams, lai sniegtu informāciju par daudzu olbaltumvielu sintēzi, kas nosaka gan ķermeņa uzbūvi, gan funkcijas. Lai veiktu olbaltumvielu sintēzi, precīza šīs informācijas kopija tiek nosūtīta ribosomām Messenger RNS formā. Dekodēšanā un sintēzē papildus i-RNS ir iesaistīts liels skaits dažādu transporta ribonukleīnskābju (t-RNS) molekulu, ribosomu un virkne enzīmu. Katra no 20 aminoskābēm saistās ar T-RNS - molekulu ar molekulu. Katrai no 20 aminoskābēm ir sava t-RNS. T-RNS ir ķīmiskas grupas, kas spēj "atpazīt" savas aminoskābes, izvēloties to no pieejamajām aminoskābēm. Tas notiek ar īpašu enzīmu palīdzību. Atzīstot tās aminoskābi, t-RNS ar to nonāk savienojumā. I-RNS ķēdes (molekulas) sākumam ir pievienota ribosoma, kas, virzoties gar i-RNS, savstarpēji savieno tieši tās aminoskābes, kuru secību kodē šīs I-RNS nukleotīdu secība. Rezultātā veidojas olbaltumvielu molekula, kuras sastāvs ir kodēts vienā no gēniem.
Nukoli- neatņemama serdes struktūras sastāvdaļa. Tie ir sfēriski ķermeņi. Tie ir ļoti maināmi, maina savu formu un struktūru, parādās un pazūd. Ir viens, divi. Katram augam noteikts skaits. Kodoli pazūd, kad šūna gatavojas dalīšanai, un pēc tam atkal parādās; acīmredzot tie ir iesaistīti ribonukleīnskābju sintēzē. Ja kodolu iznīcina ar koncentrētu rentgena vai ultravioleto staru staru, tad šūnu dalīšana tiek nomākta.
Kodola loma šūnu dzīvē. Kodols kalpo kā šūnas kontrolējošais centrs - tas vada šūnu darbību un satur iedzimtības nesējus (gēnus), kas nosaka dotā organisma īpašības. Kodola lomu var noteikt, ja, izmantojot mikroķirurģijas metodes, izņem to no šūnas un novēro tā sekas. Hemmerlings ar vienšūnu zaļajām aļģēm Acetobularia veica virkni eksperimentu, kas pierādīja svarīgu lomu šūnu augšanas regulēšanā. Šīs jūraszāles sasniedz 5 cm augstumu, izskatās kā sēne, tām ir “sakņu” un “kāju” līdzība. Augšpusē ar lielu diska formas "cepuri". Šīs aļģu šūnā ir viens kodols, kas atrodas šūnas pamatdaļā.
Hammerlings atklāja, ka, ja kāja tiek sagriezta, apakšējā daļa turpina dzīvot un vāciņš pēc operācijas ir pilnībā reģenerēts. Augšējā daļa bez kodola kādu laiku izdzīvo, bet, visbeidzot, iet bojā, nespējot atjaunot apakšējo daļu. Līdz ar to acetobularia kodols ir nepieciešams metabolisma reakcijām, kas ir augšanas pamatā.
Kodols veicina šūnu sienas veidošanos. To var parādīt eksperimentos ar aļģēm Voucheria un Spyrogyra. Atbrīvojot ūdenī šūnu saturu no sagrieztiem pavedieniem, mēs varam iegūt citoplazmas gabalus ar vienu, ar vairākiem kodoliem un bez kodoliem. Pirmajos divos gadījumos šūnu membrāna veidojās normāli. Ja nav serdes, apvalks neveidojās.
I. I. Gerasimova (1890) eksperimentos ar spirogyra tika atklāts, ka šūnas ar dubultu kodolu divkāršo hloroplasta garumu un biezumu. Bez šūnām, kurās nav kodolu, notiek fotosintēzes process, veidojas asimilācijas ciete, bet tās hidrolīzes process ir novājināts, kas izskaidrojams ar to, ka nav hidrolītisku enzīmu, kurus ribosomās var sintezēt tikai saskaņā ar kodola DNS. Protoplastu mūžs bez serdes ir zemāks un īslaicīgs. Eksperimentos I.I. Gerasimova bez kodola spirogyra šūnas dzīvoja 42 dienas un nomira. Viena no svarīgākajām kodola funkcijām ir citoplazmas apgādāšana ar ribonukleīnskābi, kas nepieciešama olbaltumvielu sintēzei šūnā. Kodola noņemšana no šūnas noved pie pakāpeniskas RNS satura samazināšanās citoplazmā un olbaltumvielu sintēzes palēnināšanās tajā.
Kodola vissvarīgākā loma ir īpašību pārnešanā no šūnas uz šūnu, no organisma uz organismu, un tas notiek kodolskaldīšanās un visas šūnas procesā.
Šūnu dalīšana.Šūnas reizina ar dalījumu. Šajā gadījumā no vienas šūnas tiek veidotas divas meitas šūnas ar tādu pašu iedzimtā materiāla komplektu, kas norobežots hromosomās kā mātes šūna. Somatiskajās šūnās hromosomas attēlo divas, tā saucamās homologās hromosomas, kurās ir iestiprināti alēlie gēni (pretēju rakstzīmju nesēji, piemēram, balto un sarkano zirņu ziedlapiņas utt.), Divu vecāku pāru pazīmes. Šajā sakarā augu ķermeņa somatiskajās šūnās vienmēr ir divkāršots hromosu komplekts, apzīmēts ar 2p. Hromosomām ir izteikta personība. Katrai sugai raksturīga iezīme ir hromosomu daudzums un kvalitāte. Tātad, zemeņu šūnās diploīdais hromosomu komplekts ir 14, (2n), ābele ir 34, topinambūrs ir 102 utt.
Mitoze (kariokinēze) - somatisko šūnu dalīšanos vispirms aprakstīja E. Russovs (1872) un I. D. Čistjakovs (1874). Tās būtība slēpjas faktā, ka daloties, no mātes šūnas tiek veidotas divas meitas šūnas ar vienādu hromosomu komplektu.Šūnu cikls sastāv no īstām starpfāzēm un mitozes. Mikroautoradiogrāfijas metode atklāja, ka garākais un grūtākais ir starpfāze - "miera" kodola periods, jo šajā laika posmā notiek kodolmateriālu dubultošanās. Starpfāze ir sadalīta trīs fāzēs:
Q1 - presintētiska (tās ilgums ir 4-6 stundas);
S - sintētisks (10-20 stundas);
Q2 - pēcsintētiskais (2–5 stundas).
Q1 fāzes laikā notiek sagatavošanās DNS samazināšanai. Un S fāzē samazinās DNS, šūna dubulto DNS piegādi. Q2 fāzē veidojas enzīmi un struktūras, kas nepieciešami mitozes ierosināšanai. Tādējādi starpfāzē DNS molekulas hromosomās tiek sadalītas divās identiskās daļās, un uz to matricas ir samontēta informācijas RNS. Pēdējais nes informāciju par specifisko olbaltumvielu struktūru citoplazmā, un kodolā katrs no DNS virzieniem aizpilda savas molekulas trūkstošo pusi. Šajā divkāršošanās (samazināšanas) procesā izpaužas unikāla DNS īpašība, kas sastāv no DNS spējas precīzi reproducēt sevi. Iegūtās meitas DNS molekulas automātiski iegūst ar precīzām sākotnējās molekulas kopijām, jo reducēšanās laikā katrai pusei pievienojas papildinošās (АТ; Г-Ц; uc) bāzes no vides.
Mitotiskās dalīšanās fāzē kļūst pamanāmas dubultās hromosomas. Metafāzē tie visi atrodas ekvatoriālajā zonā, kas atrodas vienā rindā. Veido vārpstas pavedienus (no mikrotubulēm, kas savieno viena ar otru). Kodola apvalks un nukleols pazūd. Sabiezinātas hromosomas tiek sadalītas gareniski divās meitas hromosomās. Tāda ir mitozes būtība. Tas nodrošina precīzu divkāršoto DNS molekulu sadalījumu starp meitas šūnām. Tas nodrošina iedzimtas informācijas pārraidi, kas šifrēta DNS.
Anafāzē meitas hromosomas sāk pārvietoties uz pretējiem poliem. Centrā parādās pirmie šūnu membrānas fragmenti (fragmoblasts).
Teofāzē meitas šūnās veidojas kodoli. Mātes šūnas (organelle) saturs tiek sadalīts starp iegūtajām meitas šūnām. Šūnas membrāna ir pilnībā izveidota. Tas pabeidz citokinēzi (17. att.).
Meioze - samazināšanas nodaļa tika atklāts un aprakstīts pagājušā gadsimta 90. gados V. I. Beljajevs. Dalīšanas būtība ir tāda, ka no somatiskās šūnas, kurā ir 2n (dubultā, diploīdā) hromosu kopa, veidojas četras haploīdas šūnas ar "n" - pusi no hromosu kopas. Šis dalīšanas veids ir sarežģīts un sastāv no diviem posmiem. Pirmais ir hromosomu samazināšana. Divkāršās hromosomas atrodas ekvatoriālajā zonā pa pāriem (divas paralēlas homologās hromosomas). Šajā brīdī var notikt hromosozes konjugācija (adhēzija), šķērsošana (krustošanās) un rezultātā hromosozes vietu apmaiņa. Tā rezultātā daļa tēva hromosomu gēnu kļūst par mātes hromosu daļu un otrādi. Abu hromosomu izskats tā rezultātā nemainās, bet to kvalitatīvais sastāvs kļūst atšķirīgs. Paternitātes un mātes iedzimtība tiek pārdalīta un sajaukta.
Meiozes anafāzē homologās hromosomas ar vārpstas pavedienu palīdzību atdalās pie poliem, pie kuriem pēc īsa atpūtas perioda (pavedieni pazūd, bet starpsiena starp jaunajiem kodoliem neveidojas) sākas mitozes process - metafāze, kurā visas hromosomas atrodas vienā plaknē un notiek to sadalīšana garenvirzienā. uz meitas hromosomām. Ar mitozes anafāzi ar vārpstas palīdzību tie novirzās pie poliem, kur veidojas četri kodoli un rezultātā četras haploīdas šūnas. Dažu audu šūnās to attīstības laikā noteiktu faktoru ietekmē notiek nepilnīga mitoze un hromosomu skaits kodolos dubultojas sakarā ar to, ka tie neatšķiras no poliem. Šādu dabiska vai mākslīga rakstura pārkāpumu rezultātā rodas tetraploīdi un poliploīdi. Ar mejozes palīdzību veidojas dzimumšūnas - gametas, kā arī sporas, augu seksuālās un aseksuālās reprodukcijas elementi (18. att.).
Amitoze ir tieša kodola skaldīšana. Ar amitozi neveidojas dalīšanas vārpsta, un kodola apvalks nesadalās, kā mitozē. Iepriekš amitoze tika uzskatīta par primitīvu dalīšanas formu. Tagad ir noskaidrots, ka tas ir saistīts ar ķermeņa noārdīšanos. Tā ir sarežģītāka kodola skaldīšanas vienkāršota versija. Amitoze rodas nucellus, endospermas, bumbuļu parenhīmas, lapu petioles utt. Šūnās un audos.
Mitohondriji - Tā ir spirālveida, apaļa, iegarena vai sazarota organelle.
Mitohondriju jēdzienu pirmo reizi ierosināja Benda 1897. gadā. Mitohondrijus dzīvajās šūnās var noteikt, izmantojot fāzu kontrastu un traucējumu mikroskopiju graudu, granulu vai pavedienu formā. Tās ir diezgan mobilas struktūras, kuras var pārvietoties, saplūst viena ar otru, dalīties. Ja ar īpašām metodēm krāso mirušajās šūnās gaismas mikroskopijā, mitohondriji izskatās kā mazi graudi (granulas), kas difūziski izkliedēti citoplazmā vai koncentrēti dažās specifiskās zonās.
Glikozes un tauku iznīcināšanas rezultātā skābekļa klātbūtnē mitohondrijos tiek ģenerēta enerģija, organiskās vielas tiek pārveidotas ūdenī un oglekļa dioksīdā. Tādā veidā dzīvnieku organismi iegūst dzīvībai nepieciešamo enerģiju. Enerģija tiek uzkrāta adenozīna trifosfātā (ATP), precīzāk, tā makroerģiskajās saitēs. Mitohondriju funkcija ir cieši saistīta ar organisko savienojumu oksidēšanu un to sadalīšanās laikā atbrīvotās enerģijas izmantošanu ATP molekulu sintēzei. Tāpēc mitohondrijus bieži sauc par šūnu enerģijas stacijām vai šūnu elpošanas organelliem. ATP darbojas kā enerģijas piegādātājs, pārnesot vienu no ar enerģiju bagātajām termināla fosfātu grupām uz citu molekulu un pārvēršas ADP.
Tiek uzskatīts, ka evolūcijā mitohondriji bija prokarioti mikroorganismi, kas senās šūnas ķermenī kļuva par simbiotikām. Pēc tam tie kļuva ļoti nepieciešami, kas bija saistīts ar skābekļa satura palielināšanos Zemes atmosfērā. No vienas puses, mitohondriji no šūnas izņēma lieko toksisko skābekli, no otras puses, tas sniedza enerģiju.
Bez mitohondrijiem šūna praktiski nespēj izmantot skābekli kā vielu, kas nodrošina enerģijas piegādi, un savas enerģijas vajadzības var apmierināt tikai caur anaerobiem procesiem. Tādējādi skābeklis ir inde, bet inde ir ļoti svarīga šūnai, un skābekļa pārpalikums ir tikpat kaitīgs kā tā deficīts.
Mitohondriji var mainīt savu formu un pārcelties uz tām šūnas vietām, kur nepieciešamība pēc tām ir visaugstākā. Tātad kardiomiocītos mitohondriji atrodas netālu no miofibrilliem, nieru kanāliņu šūnās pie bazālajiem izvirzījumiem utt. Šūnā ir līdz tūkstoš mitohondriju, un to skaits ir atkarīgs no šūnas aktivitātes.
Mitohondriju vidējais šķērsvirziena izmērs ir 0,5 ... 3 mikroni. Atkarībā no lieluma izšķir mazas, vidējas, lielas un milzu mitohondrijas (tās veido sazarotu tīklu - mitohondriju retikulumu). Mitohondriju lielums un skaits ir cieši saistīti ar šūnas darbību un tās enerģijas patēriņu. Tie ir ārkārtīgi mainīgi un atkarībā no šūnu aktivitātes, skābekļa satura, hormonālās ietekmes, var uzbriest, mainīt cristae skaitu un struktūru, mainīties pēc skaita, formas un lieluma, kā arī fermentatīvās aktivitātes.
Mitohondriju masas blīvums, to iekšējās virsmas attīstības pakāpe un citi rādītāji ir atkarīgi no šūnas enerģijas vajadzībām. Limfocītos ir tikai dažas mitohondrijas, un aknu šūnās - 2 ... 3 tūkstoši.
Mitohondriji sastāv no matricas, iekšējās membrānas, perimitohondriju telpas un ārējās membrānas. Ārējā mitohondriju membrāna atdala organeli no hialoplazmas. Parasti tam ir gludas kontūras un tas ir aizvērts tā, lai tas būtu membrānas maisiņš.
Ārējo membrānu no iekšējās atdala perimitohondrijas telpa ar platumu aptuveni 10 ... 20 nm. Iekšējā mitohondriju membrāna ierobežo mitohondriju - matricas - faktisko iekšējo saturu. Iekšējā membrāna veido daudzus izvirzījumus uz iekšu no mitohondrijiem, kuriem ir plakanas grēdas vai čaumalas.
Pēc formas kraukļi var būt plākšņu (trabekulāri) un kanāliņu (griezumā multivikulāri) formā, un tie ir vērsti gareniski vai šķērsām attiecībā uz mitohondrijiem.
Katru mitohondriju piepilda matrica, kas elektronu mikrogrāfijās izskatās blīvāka nekā apkārtējā citoplazma. Mitohondriju matrica ir viendabīga (viendabīga), dažreiz smalkgraudaina, ar dažādu elektronu blīvumu. Tajā atklāti plāni pavedieni ar biezumu aptuveni 2 ... 3 nm un granulas ar izmēru aptuveni 15 ... 20 nm. Matricas šķipsnas ir DNS molekulas, un mazas granulas ir mitohondriju ribosomas. Matricā ir fermenti, viena vienšūna, cikliska DNS, mitohondriju ribosomas, daudzi Ca 2+ joni.
Mitohondriju olbaltumvielu sintēzes autonomo sistēmu attēlo DNS molekulas, kas nesatur histonu. DNS ir īsa, tai ir gredzena (cikliska) forma un tajā ir 37 gēni. Atšķirībā no kodola DNS, tajā praktiski nav nekodētu nukleotīdu secību. Struktūras un organizācijas pazīmes apvieno mitohondriju DNS ar baktēriju šūnu DNS. Mitohondriju DNS notiek dažāda veida RNS molekulu sintēze: informācija, pārnešana (transports) un ribosomāli. Informācija mitohondriju RNS netiek pakļauta splicēšanai (griešanas sekcijas, kurām nav informācijas slodzes). Mitohondriju DNS molekulu mazais izmērs nevar noteikt visu mitohondriju olbaltumvielu sintēzi. Lielākā daļa mitohondriju olbaltumvielu atrodas šūnas kodola ģenētiskā kontrolē un tiek sintezētas citoplazmā, jo mitohondriju DNS ir vāji ekspresēts un var nodrošināt tikai daļu no oksidatīvās fosforilēšanās ķēdes fermentiem. Mitohondriju DNS kodē ne vairāk kā desmit olbaltumvielas, kas ir lokalizētas membrānās un ir strukturālās olbaltumvielas, kas ir atbildīgas par mitohondriju membrānu atsevišķu funkcionālu olbaltumvielu kompleksu pareizu integrāciju. Tiek sintezēti arī olbaltumvielas, kas veic transporta funkcijas. Šāda olbaltumvielu sintēzes sistēma nenodrošina visas mitohondriju funkcijas, tāpēc mitohondriju autonomija ir ierobežota un relatīva.
Zīdītājiem mitohondrijas apaugļošanās laikā tiek pārnestas tikai caur olu, un spermatozoīdi jaunajā organismā ievada nukleīnu DNS.
Mitohondriju matricā veidojas ribosomas, kas atšķiras no citoplazmatiskajām ribosomām. Viņi piedalās vairāku mitohondriju olbaltumvielu sintēzē, kuras kodols nav kodēts. Mitohondriju ribosomu sedimentācijas skaits ir 60 (atšķirībā no citoplazmas, kur sedimentācijas skaits ir 80). Sedimentācijas numurs ir nogulsnēšanās ātrums centrifugēšanas un ultracentrifugēšanas laikā. Pēc struktūras mitohondriju ribosomas ir tuvu prokariotu organismu ribosomām, bet ir mazākas un ir jutīgas pret noteiktām antibiotikām (hloramfenikolu, tetraciklīnu utt.).
Iekšējā mitohondriju membrānā ir augsta selektivitātes pakāpe vielu pārvadāšanā. Oksidējošās fosforilēšanas ķēdes enzīmi, elektronu nesējproteīni, ATP, ADP, piruvāts un citas transportsistēmas, kas atrodas cieši blakus viena otrai, ir piestiprinātas pie tās iekšējās virsmas.Sakarā ar fermentu ciešu izvietojumu uz iekšējās membrānas tiek nodrošināta augsta bioķīmisko procesu konjugācija (savstarpēja savienošanās), kas palielina ātrumu un katalītisko procesu efektivitāte.
Elektronu mikroskopija atklāj sēņu daļiņas, kas izvirzās matricas lūmenā. Viņiem ir ATP sintētiskā (veido ATP no ADP) aktivitāte. Elektronu transportēšana notiek pa elpošanas ķēdi, lokalizējoties iekšējā membrānā, kurā ir četri lieli enzīmu kompleksi (citohromi). Kad elektroni iziet caur elpošanas ķēdi, ūdeņraža joni no matricas tiek izsūknēti perimitohondriju telpā, kas nodrošina protonu gradienta (sūkņa) veidošanos. Šī gradienta enerģija (vielu koncentrācijas atšķirības un membrānas potenciāla veidošanās) tiek izmantota ATP sintēzei un metabolītu un neorganisko jonu transportēšanai. Iekšējā membrānā esošie nesējproteīni caur to transportē organiskos fosfātus, ATP, ADP, aminoskābes, taukskābes, tri- un dikarbonskābes.
Mitohondriju ārējā membrāna ir labāk caurlaidīga vielām ar zemu molekulmasu, jo tai ir daudz hidrofilu olbaltumvielu kanālu. Uz ārējās membrānas atrodas specifiski receptoru kompleksi, caur kuriem olbaltumvielas no matricas tiek transportētas perimitohondriju telpā.
Ķīmiskā sastāva un īpašību ziņā ārējā membrāna ir tuvu citām intracelulārajām membrānām un plazmolemmām. Tas satur fermentus, kas metabolizē taukus, aktivizē (katalizē) aminoskābju, aminoskābes pārvēršanu. Ja ārējās membrānas fermenti paliek aktīvi, tad tas ir mitohondriju funkcionālās drošības indikators.
Mitohondrijos ir divi autonomi apakšnodaļas. Tā kā perimitochondrial telpa jeb ārējā mitohondriju kamera (ārējais apakšnodalījums) veidojas hialoplazmas olbaltumvielu kompleksu iespiešanās dēļ, mitohondriju DNS sintētiskās aktivitātes dēļ daļēji veidojas iekšējais apakšnodalījums (mitohondriju matrica). Iekšējais apakšnodaļa (matrica) satur DNS, RNS un ribosomas. Tam ir augsts Ca 2+ jonu līmenis, salīdzinot ar hialoplazmu. Ārējā apakšnodalījumā uzkrājas ūdeņraža joni. Ārējo un iekšējo apakšsekciju fermentatīvā aktivitāte, olbaltumvielu sastāvs ir ļoti atšķirīgs. Iekšējam apakšnodalījumam ir lielāks elektronu blīvums nekā ārējam.
Specifiski mitohondriju marķieri ir citohroma oksidāzes un sukcinātu dehidrogenāzes enzīmi, kuru noteikšana ļauj kvantitatīvi raksturot enerģijas procesus mitohondrijos.
Mitohondriju galvenā funkcija - ATP sintēze. Sākumā hialoplazmā cukuri (glikoze) tiek iznīcināti līdz pienskābēm un piruvavām (piruvāts), vienlaikus sintezējot nelielu daudzumu ATP. Vienas glikozes molekulas glikolīzes rezultātā tiek izmantotas divas ATP molekulas, no kurām veidojas četras. Tādējādi tikai divas ATP molekulas veido pozitīvo bilanci. Šie procesi notiek bez skābekļa (anaerobā glikolīze).
Visi nākamie enerģijas ražošanas posmi notiek aerobās oksidācijas procesā, kas nodrošina liela daudzuma ATP sintēzi. Šajā gadījumā organiskās vielas iznīcina līdz CO 2 un ūdenim. Oksidēšanu papildina protonu nodošana to akceptoriem. Šīs reakcijas tiek veiktas, izmantojot vairākus trikarbonskābes cikla enzīmus, kas atrodas mitohondriju matricā.
Kristāla membrānās ir iebūvētas elektronu pārneses sistēmas un ar tām saistītā ADP fosforilēšana (oksidatīvā fosforilēšana). Šajā gadījumā notiek elektronu pārnešana no viena elektronu akceptora olbaltumvielas uz otru un, visbeidzot, to saistīšanās ar skābekli, kā rezultātā veidojas ūdens. Tajā pašā laikā daļa enerģijas, kas izdalās šādas oksidācijas laikā elektronu pārneses ķēdē, tiek uzglabāta makroerģiskas saites formā ADP fosforilēšanas laikā, kas noved pie liela skaita ATP molekulu veidošanās - galvenā starpšūnu enerģijas ekvivalenta. Oksidatīvās fosforilēšanās process notiek uz mitohondriju cristae membrānām, izmantojot oksidācijas ķēdes proteīnus un ADP ATP sintetāzes fosforilēšanas fermentu, kas atrodas šeit. Oksidējošās fosforilēšanas rezultātā no vienas glikozes molekulas veidojas 36 ATP molekulas.
Dažiem hormoniem un vielām uz mitohondrijiem ir īpaši (afinitātes) receptori. Trijodtironīns parasti paātrina mitohondriju sintētisko aktivitāti. Interleikīns-1 un augsta trijodtironīna koncentrācija atdala oksidatīvās fosforilēšanās ķēdes, izraisot mitohondriju pietūkumu, ko papildina siltumenerģijas veidošanās palielināšanās.
Jaunas mitohondrijas veidojas daloties, sašaurinoties vai veidojot pumpurus. Pēdējā gadījumā veidojas protomitohondriji, pakāpeniski palielinot izmēru.
Protomitochondria ir maza organelle ar ārējo un iekšējo membrānu. Iekšējā membrānā nav vai ir mazattīstīti kraukļi. Organellu raksturo zems aerobās fosforilēšanās līmenis. Ar sašaurināšanās veidošanos mitohondriju saturs tiek sadalīts starp diviem jauniem diezgan lieliem organelliem. Izmantojot jebkuru reprodukcijas metodi, katram no jaunizveidotajiem mitohondrijiem ir savs genoms.
Vecākus mitohondrijus iznīcina autolīze (šūnas pašsagremošana, izmantojot lizosomas), veidojot autolizosomas. No autolizosomas veidojas atlikuma ķermenis. Pēc pilnīgas sagremošanas ķermeņa atlikuma saturs, kas sastāv no organiskām vielām ar mazu molekulmasu, izdalās ar eksocitozi. Ar nepilnīgu gremošanu mitohondriju paliekas var uzkrāties šūnā slāņveida ķermeņu vai granulu veidā ar nipofuscīnu. Nešķīstošie kalcija sāļi uzkrājas mitohondriju daļā ar kristālu veidošanos - pārkaļķošanos. Mitohondriju deģenerācijas produktu uzkrāšanās var izraisīt šūnu distrofiju.
Mitohondriju struktūra un funkcijas ir diezgan sarežģīts jautājums. Organelle klātbūtne ir raksturīga gandrīz visiem kodolorganismiem - gan autotrofiem (augiem, kas spēj fotosintēzi), gan heterotrofiem, kas ir gandrīz visi dzīvnieki, daži augi un sēnītes.
Mitohondriju galvenais mērķis ir organisko vielu oksidēšana un sekojoša enerģijas izmantošana, kas izdalās šī procesa rezultātā. Šī iemesla dēļ organellām ir arī otrs (neoficiāls) nosaukums - šūnu enerģijas stacijas. Tos dažreiz sauc par "katabolisma plastidiem".
Kas ir mitohondriji
Šis termins ir radies grieķu valodā. Tulkojumā šis vārds nozīmē "pavediens" (mitos), "sēkla" (hondrions). Mitohondriji ir pastāvīgi organoīdi, kuriem ir liela nozīme normālai šūnu darbībai un tie ļauj eksistēt visam organismam.

"Stacijām" ir īpaša iekšējā struktūra, kas mainās atkarībā no mitohondriju funkcionālā stāvokļa. To forma var būt divu veidu - ovāla vai iegarena. Pēdējam bieži ir sazarots izskats. Organellu skaits vienā šūnā svārstās no 150 līdz 1500.
Īpašs gadījums ir dzimumšūnas. Spermā ir tikai viena spirālveida organelle, savukārt sieviešu dzimuma gametas satur simtiem tūkstošu vairāk mitohondriju. Šūnā organellas nav fiksētas vienā vietā, bet var pārvietoties gar citoplazmu, apvienoties viena ar otru. To izmērs ir 0,5 mikroni, garums var sasniegt 60 mikronus, bet minimālais skaitlis ir 7 mikroni.

Vienas “elektrostacijas” lieluma noteikšana nav viegls uzdevums. Fakts ir tāds, ka, pārbaudot elektronu mikroskopā, šķēlēs nokļūst tikai daļa no organelliem. Tā notiek, ka spirālveida mitohondrijās ir vairākas sadaļas, kuras var izmantot atsevišķām, neatkarīgām struktūrām.
Tikai trīsdimensiju attēls ļaus mums noskaidrot precīzu šūnu struktūru un saprast, vai mēs runājam par 2–5 atsevišķiem organoīdiem vai par tiem, kuriem ir sarežģīta mitohondriju forma.
Strukturālās iezīmes
Mitohondriju membrāna sastāv no diviem slāņiem: ārējā un iekšējā. Pēdējā ietilpst dažādi izaugumi un krokas, kurām ir lapveidīga un cauruļveida forma.
Katrai membrānai ir īpašs ķīmiskais sastāvs, noteikts daudzums noteiktu enzīmu un noteikts mērķis. Ārējo apvalku no iekšpuses atdala ar starpmembrānu, kuras biezums ir 10-20 nm.
Orgānu struktūra attēlā ar parakstiem izskatās ļoti skaidra.

Mitohondriju struktūra
Apskatot struktūras diagrammu, var izveidot šādu aprakstu. Viskozo telpu mitohondriju iekšienē sauc par matricu. Tās sastāvs rada labvēlīgu vidi nepieciešamo ķīmisko procesu plūsmai tajā. Tas satur mikroskopiskas granulas, kas veicina reakcijas un bioķīmiskos procesus (piemēram, tie uzkrāj glikogēna jonus un citas vielas).
Matrica satur DNS, koenzīmus, ribosomas, t-RNS, neorganiskos jonus. ATP sintāze un citohromi atrodas uz apvalka iekšējā slāņa virsmas. Fermenti veicina tādus procesus kā Krebsa cikls (CTK), oksidatīvā fosforilēšanās utt.
Tādējādi galveno organoīda uzdevumu veic gan matrica, gan apvalka iekšējā puse.
Mitohondriju funkcija
"Elektrostaciju" mērķi var raksturot ar diviem galvenajiem uzdevumiem:
- enerģijas ražošana: tajos tiek veikti oksidācijas procesi ar sekojošu ATP molekulu izdalīšanos;
- ģenētiskās informācijas glabāšana;
- dalība hormonu, aminoskābju un citu struktūru sintēzē.
Oksidācijas un enerģijas ražošanas process notiek vairākos posmos:


ATP sintēzes shematisks rasējums
Ir vērts atzīmēt: Krebsa cikla (citronskābes cikla) \u200b\u200brezultātā ATP molekulas neveidojas, molekulas tiek oksidētas un izdalās oglekļa dioksīds. Šis ir starpposms starp glikolīzi un elektronu transporta ķēdi.
Tabula "Mitohondriju funkcijas un struktūra"

Kas nosaka mitohondriju skaitu šūnā
Dominējošais organoīdu skaits uzkrājas blakus tām šūnas daļām, kur ir nepieciešami enerģijas resursi. Jo īpaši miofibrilu apgabalā tiek savākts liels daudzums organoļu, kas ir daļa no muskuļu šūnām, kas nodrošina to saraušanos.

Vīriešu dzimumšūnās struktūras ir lokalizētas ap flagellum asi - tiek pieņemts, ka nepieciešamība pēc ATP ir saistīta ar nepārtrauktu gametas astes kustību. Mitohondriju izvietojums vienšūņos, kuru pārvietošanai tiek izmantotas īpašas ciliakas, izskatās, ka organellas uzkrājas zem membrānas pie pamatnes tieši tādā pašā veidā.
Kas attiecas uz nervu šūnām, mitohondriju lokalizācija tiek novērota netālu no sinapsēm, caur kurām tiek pārraidīti nervu sistēmas signāli. Šūnās, kas sintezē olbaltumvielas, ergastoplasmas zonās uzkrājas organellās - tās piegādā enerģiju, kas nodrošina šo procesu.
Kas atklāja mitohondrijus
Šūnu struktūra savu nosaukumu ieguva 1897.-1898. Gadā, pateicoties C. Brand. Otto Vāburgs 1920. gadā spēja pierādīt šūnu elpošanas saistību ar mitohondrijiem.
Secinājums
Mitohondriji ir vissvarīgākā dzīvās šūnas sastāvdaļa, kas darbojas kā enerģijas stacija, kas ražo ATP molekulas, tādējādi nodrošinot šūnu dzīvībai svarīgās aktivitātes procesus.
Mitohondriju darba pamatā ir organisko savienojumu oksidēšana, kā rezultātā rodas enerģijas potenciāls.
Ārējā membrāna
Iekšējā membrāna
Matrica m-on, matrix, krista. tai ir gludas kontūras, neveido izvirzījumus vai krokas. Tas aizņem apmēram 7% no visu šūnu membrānu laukuma. Tās biezums ir aptuveni 7 nm, tas nav savienots ar citām citoplazmas membrānām un ir noslēgts pats pret sevi, tāpēc tas ir membrānas maisiņš. Ārējā membrāna ir atdalīta no iekšējās starpposma telpa apmēram 10-20 nm plata. Iekšējā membrāna (apmēram 7 nm bieza) ierobežo faktisko mitohondriju iekšējo saturu,
tā matrica vai mitoplazma. Mitohondriju iekšējās membrānas raksturīga iezīme ir to spēja mitohondriju iekšpusē veidot daudzus izvirzījumus. Šādiem izvirzījumiem visbiežāk ir plakanas grēdas vai čaumalas. Attālums starp membrānām kristālā ir apmēram 10-20 nm. Bieži vien cristae var sazaroties vai veidot pirksta formas procesus, saliekties un tiem nav izteiktas orientācijas. Vienšūņu aļģēs, dažās augstāku augu un dzīvnieku šūnās, vienšūnu aļģēs iekšējās membrānas izaugumi izskatās kā caurules (cauruļveida čaumalas).
Mitohondriju matricai ir smalkgraudaina viendabīga struktūra, dažreiz tajā tiek uztverti plāni pavedieni, kas savākti bumbiņā (apmēram 2-3 nm) un granulas apmēram 15-20 nm. Tagad ir kļuvis zināms, ka mitohondriju matricas šķipsnas ir DNS molekulas mitohondriju nukleoīda sastāvā, un mazas granulas ir mitohondriju ribosomas.
Mitohondriju funkcija
1. Mitohondrijās notiek ATP sintēze (sk. Oksidējošo fosforilēšanu)
Starpslāņu pH ~ 4, matricas pH ~ 8 | olbaltumvielu saturs metros: 67% ir matrica, 21% ir raktuves atklājums, 6% ir raktuves iekšpuse un 6% ir raktuves
Distemper - viena mitohondriju sistēma
ārējā m-na: poru poras ļauj iziet cauri 5 kD | iekšējais m-na: kardiolipīniem necaurlaidīgs jonu jonu |
starptautiskā ražošana: fermentu grupas fosforilē nukleotīdus un nukleotīdu cukurus
Iekšzemē:
matrica: vielmaiņas fermenti - lipīdu oksidācija, ogļhidrātu oksidēšana, trikarbonskābes cikls, Krebsa cikls
Izcelsme no baktērijām: Pelomyxa palustris amēba nesatur eikariotu savienojumus, dzīvo simbiozē ar aerobajām baktērijām | pašu DNS | OX līdzīgas baktērijas
Mitohondriju DNS
Miohondriju sadalīšana
atkārtots
starpfāzē | replikācija nav saistīta ar S fāzi | cl cikla laikā mitohis vienreiz tiek sadalīts divās daļās, veidojot sašaurinājumu, vispirms sašaurinājums iekšpusē m-nav | ~ 16,5 kb | gredzens kodē 2 rRNS 22 tRNS un 13 olbaltumvielas |
olbaltumvielu transports: signāla peptīds | amfifiliskā čokurošanās | mitohondriju atpazīšanas receptori |
Oksidējošā fosforilēšana
Elektronu pārvades ķēde
ATP sintāze
aknu šūnās dzīvo ~ 20 dienas mitohondriju dalīšanās, veidojoties sašaurinājumam
16569 bp \u003d 13 olbaltumvielas, 22tRNS, 2pRNS | gluda ārējā mn (porins - olbaltumvielu caurlaidība līdz 10 kDa) salocīta iekšējā (crista) mn (75% olbaltumvielu): transportēšanas nesēja olbaltumvielas, f-you, elpošanas ķēdes un ATP sintāzes komponenti, kardiolipīna) matrica ( bagātināts ar f-tami citrāta ciklu) inter 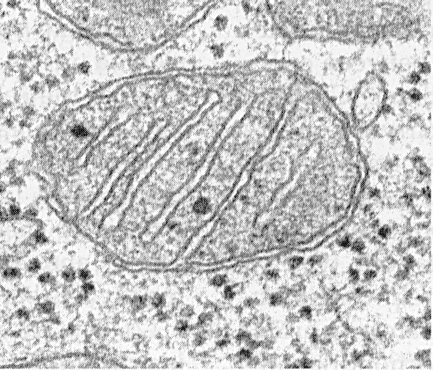
Mitohondriji vai hondriosomas (no grieķu valodas. Mitos - pavediens, chondrion - graudi, soma - ķermenis) ir granulētas vai pavedienveidīgas organellas, kas atrodas vienšūņu, augu un dzīvnieku citoplazmā. Mitohondrijus var novērot dzīvās šūnās, jo tām ir diezgan augsts blīvums. Dzīvajās šūnās mitohondriji var kustēties, kustēties, saplūst viens ar otru.
Dažādās sugās mitohondriju izmēri ir ļoti mainīgi, kā arī mainīga ir to forma (199. att.). Neskatoties uz to, lielākajā daļā šūnu šo struktūru biezums ir salīdzinoši nemainīgs (apmēram 0,5 μm), un garums svārstās, pavedienveidīgajās formās sasniedzot līdz 7–60 μm.
Mitohondriju lieluma un skaita izpēte nav tik vienkārša lieta. Tas ir saistīts ar faktu, ka mitohondriju izmērs un skaits, kas ir redzams uz ultra-plānām sekcijām, neatbilst realitātei.
Parastie aprēķini liecina, ka aknu šūnā atrodas apmēram 200 mitohondriju. Tas veido vairāk nekā 20% no kopējās citoplazmas un apmēram 30-35% no kopējā olbaltumvielu daudzuma šūnā. Visu aknu šūnu mitohondriju virsmas laukums ir 4-5 reizes lielāks nekā tās plazmas membrānas virsma. Lielākā daļa mitohondriju atrodas oocītos (apmēram 300 000) un milzu amēbas Chaos haosā (līdz 500 000).
Zaļo augu šūnās mitohondriju ir mazāk nekā dzīvnieku šūnās, jo hloroplasti var veikt daļu no savām funkcijām.
Mitohondriju lokalizācija šūnās ir atšķirīga. Parasti mitohondrijas uzkrājas netālu no tām citoplazmas vietām, kur ir nepieciešams ATP, kas veidojas mitohondrijos. Tātad skeleta muskuļos mitohondriji atrodas netālu no miofibrilām. Spermā mitohondriji veido spirāles lietu ap flagellum asi; tas, iespējams, ir saistīts ar nepieciešamību izmantot ATP, lai pārvietotu spermas asti. Līdzīgi vienšūņos un citās šūnās, kas aprīkotas ar ciliaku, mitohondrijas tiek lokalizētas tieši zem šūnu membrānas ciliaku pamatnē, kurām nepieciešams ATP. Nervu šūnu aksonos mitohondriji atrodas netālu no sinapsēm, kur notiek nervu impulsa pārnešanas process. Sekrecijas šūnās, kas sintezē lielu daudzumu olbaltumvielu, mitohondrijas ir cieši saistītas ar ergastoplasma zonām; tie, iespējams, piegādā ATP, lai aktivizētu aminoskābes un olbaltumvielu sintēzi ribosomās.
Mitohondriju ultrastruktūra.
Mitohondrijiem neatkarīgi no to lieluma vai formas ir universāla struktūra, to ultrastruktūra ir vienota. Mitohondriji ir ierobežoti ar divām membrānām (205. att.). Ārējā mitohondriju membrāna atdala to no hialoplazmas, tai ir gludas kontūras, neveido izvirzījumus vai krokas, un biezums ir aptuveni 7 nm. Tas aizņem apmēram 7% no visu šūnu membrānu laukuma. Membrāna nav savienota ar citām citoplazmas membrānām, pati ir aizvērta un ir membrānas maisiņš. Ārējo membrānu no iekšējās atdala ar starpmembrānu telpu ar platumu aptuveni 10-20 nm. Iekšējā membrāna (apmēram 7 nm bieza) ierobežo mitohondriju, tās matricas vai mitoplazmas faktisko iekšējo saturu. Mitohondriju iekšējā membrāna mitohondriju iekšpusē veido daudzus izspiejumus. Šādiem izvirzījumiem visbiežāk ir plakanas grēdas vai čaumalas.
Aknu šūnas iekšējās mitohondriju membrānas kopējā virsma ir aptuveni viena trešdaļa no visu šūnu membrānu virsmas. Sirds muskuļa šūnu mitohondrijos ir trīs reizes vairāk kraukšķu nekā aknu mitohondrijās, kas atspoguļo atšķirības dažādu šūnu mitohondriju funkcionālajās slodzēs. Attālums starp membrānām kristālā ir apmēram 10-20 nm.
Mitohondriju krītiņi, kas stiepjas no iekšējās membrānas un stiepjas matricas virzienā, pilnībā neaizbloķē mitohondriju dobumu un nepārkāpj to piepildošās matricas nepārtrauktību.
Krītes orientācija attiecībā pret mitohondriju garo asi dažādām šūnām ir atšķirīga. Orientācija var būt perpendikulāra (aknu, nieru) kraukšķis; sirds muskulī tiek novērots krustveida gareniskais izvietojums. Crista var sazaroties vai veidot pirkstu formas procesus, saliekties un tiem nav izteiktas orientācijas (208. att.). Vienšūņu aļģēs, dažās augstāku augu un dzīvnieku šūnās, vienšūnu aļģēs iekšējās membrānas izaugumi izskatās kā caurules (cauruļveida čaumalas).
Mitohondriju matricai ir smalkgraudaina viendabīga struktūra, kurā DNS molekulas tiek noteiktas plānas pavedienu veidā, kas savākti bumbiņā (apmēram 2-3 nm), un mitohondriālās ribosomas granulu formā, kuru izmērs ir aptuveni 15-20 nm. Magnija un kalcija sāļu nogulsnes matricā veido lielas (20–40 nm) blīvas granulas.
Mitohondriju funkcijas.
Mitohondriji veic ATP sintēzi, kas notiek organisko substrātu oksidācijas un ADP fosforilēšanas procesu rezultātā.
Sākotnējos ogļhidrātu oksidācijas posmus sauc par anaerobo oksidāciju vai glikolīze un rodas hialoplazmā un neprasa skābekļa piedalīšanos. Anaerobās enerģijas ražošanas laikā oksidācijas substrāts ir heksozes un, pirmkārt, glikoze; dažām baktērijām ir spēja iegūt enerģiju, oksidējot pentozes, taukskābes vai aminoskābes.
Glikozes potenciālās enerģijas daudzums, kas atrodas saiknēs starp C, H un O atomiem, ir aptuveni 680 kcal uz molu (t.i., 180 g glikozes).
Dzīvā šūnā šis milzīgais enerģijas daudzums tiek atbrīvots pakāpeniska procesa veidā, ko kontrolē vairāki oksidējoši fermenti, un tas nav saistīts ar ķīmiskās saites enerģijas pāreju siltumā, kā sadegšanas laikā, bet pārvēršas par makroenerģētisko saiti ATP molekulās, kuras tiek sintezētas, izmantojot atbrīvoto enerģiju no ADP un fosfāts.
Triozes, kas veidojas glikolīzes rezultātā, galvenokārt pirūdskābe, ir iesaistītas tālākā oksidācijā mitohondrijos. Šajā gadījumā notiek visu ķīmisko saišu šķelšanās enerģijas izmantošana, kas izraisa CO 2 izdalīšanos, skābekļa patēriņu un liela daudzuma ATP sintēzi. Šie procesi ir saistīti ar trikarbonskābju oksidācijas ciklu un ar elektronu pārnešanas elpošanas ķēdi, kur notiek ADP fosforilēšanās un šūnu “kurināmā” un ATP molekulu sintēze (209. att.).
Trikarbonskābes ciklā (Krebsa ciklā vai citronskābes ciklā) piruvāts, kas veidojas glikolīzes rezultātā, vispirms zaudē CO 2 molekulu un, oksidējoties par acetātu (divu oglekļa savienojumu), apvienojas ar koenzīmu A. Pēc tam acetilkoenzīms A apvienojas ar oksalacetātu (četru oglekļa savienojumu), veido sešu oglekļa citrātu (citronskābi). Pēc tam notiek šī sešu oglekļa savienojuma oksidēšanas cikls līdz četru oglekļa oksalacetātam, atkal saistoties ar acetilkoenzīmu A, un pēc tam cikls atkārtojas. Šīs oksidācijas laikā izdalās divas CO 2 molekulas, un oksidācijas laikā atbrīvotie elektroni tiek pārnesti uz akceptoru koenzīma molekulām (NAD-nikotinamīda adenīna dinukleotīds), kas tos vēl vairāk iesaista elektronu pārneses ķēdē. Tāpēc trikarbonskābju ciklā nav pašas ATP sintēzes, bet notiek molekulu oksidēšana, elektronu pārnešana uz akceptoriem un CO 2 izmešana. Visi iepriekš aprakstītie notikumi mitohondrijās notiek to matricā.
Sākotnējā substrāta oksidēšana noved pie CO 2 un ūdens izdalīšanās, bet siltumenerģija netiek atbrīvota, kā sadegšanas laikā, bet veidojas ATP molekulas. Tos sintezē cita veida olbaltumvielas, kas nav tieši saistītas ar oksidāciju. Iekšējās mitohondriju membrānās uz membrānu virsmas, kas ieskatās matricā, atrodas lieli olbaltumvielu kompleksi, fermenti, ATP sintetāzes. Elektronu mikroskopā tie ir redzami tā saukto “sēņu” ķermeņu formā, kas pilnībā izlīdzina membrānu virsmu, ieskatoties matricā. Ķermenim ir kāja un galva ar diametru 8-9 nm. Līdz ar to gan oksidācijas ķēdes, gan ATP sintēzes enzīmi tiek lokalizēti iekšējās mitohondriju membrānās (201.b att.).
Elpošanas ķēde ir galvenā enerģijas pārveidošanas sistēma mitohondrijos. Šeit notiek elpošanas ķēdes elementu secīga oksidēšana un atjaunošana, kā rezultātā enerģija tiek atbrīvota nelielās porcijās. Šīs enerģijas dēļ trīs ķēdes punktos ATP veidojas no ADP un fosfāta. Tāpēc tiek teikts, ka oksidēšanās (elektronu pārnešana) ir saistīta ar fosforilēšanu (ADP + Fn → ATP, t.i., notiek oksidatīvā fosforilēšanās.
Elektronu transportēšanas laikā atbrīvotā enerģija tiek glabāta protonu gradienta veidā uz membrānas. Izrādījās, ka elektronu pārnešanas laikā mitohondriju membrānā katrs elpošanas ķēdes komplekss novirza brīvo oksidācijas enerģiju, lai pārvietotu protonus (pozitīvos lādiņus) caur membrānu, no matricas uz starpmembranālo telpu, kas noved pie membrānas potenciālo atšķirību veidošanās: starpmembranālajā telpā dominē pozitīvas lādiņas, un negatīvs - no mitohondriju matricas puses. Kad ir sasniegta potenciālā starpība (220 mV), ATP sintetāzes olbaltumvielu komplekss sāk transportēt protonus atpakaļ uz matricu, vienlaikus pārvēršot vienu enerģijas veidu citā: tas veido ATP no ADP un neorganisko fosfātu. Tas ir oksidējošu procesu un sintētisku procesu apvienojums ar ADP fosforilēšanu. Kamēr substrāti oksidējas, kamēr protoni tiek iesūknēti caur iekšējo mitohondriju membrānu, notiek ar to saistītā ATP sintēze, t.i. notiek oksidatīvā fosforilēšanās.
Šos divus procesus var atdalīt. Šajā gadījumā turpinās elektronu pārnešana, tāpat kā substrāta oksidēšana, bet ATP sintēze nenotiek. Šajā gadījumā oksidācijas laikā atbrīvotā enerģija tiek pārveidota par siltumenerģiju.
Oksidējošā fosforilēšanās baktērijās.
Prokariotu šūnās, kas spēj oksidatīvi fosforilēties, trikarbonskābes cikla elementi tiek lokalizēti tieši citoplazmā, un elpošanas ķēde un fosforilēšanās fermenti ir saistīti ar šūnu membrānu, tās izvirzījumiem izvirzoties citoplazmā, ar tā saucamajām mezosomām (212. att.). Jāatzīmē, ka šādas baktēriju mezosomas var saistīt ne tikai ar aerobās elpošanas procesiem, bet arī dažās sugās piedalās šūnu dalīšanā, DNS izplatīšanas procesā jaunajās šūnās, šūnas sienas veidošanā utt.
Uz plazmas membrānas dažu baktēriju mezosomās tiek veikti gan oksidācijas, gan ATP sintēzes konjugētie procesi. Sfēriskas daļiņas, kas līdzīgas tām, kas atrodamas eikariotu šūnu mitohondrijos, tika atrasti elektronu mikroskopā baktēriju plazmas membrānu frakcijās. Tādējādi baktēriju šūnās, kas spēj oksidatīvi fosforilēties, plazmas membrānai ir līdzīga loma kā eikariotu šūnu iekšējai mitohondrijai.
Mitohondriju skaita palielināšanās.
Mitohondriji var palielināt to skaitu, jo īpaši, daloties šūnās vai palielinoties šūnas funkcionālajai slodzei. Pastāvīga mitohondriju atjaunošanās. Piemēram, aknās mitohondriju vidējais dzīves ilgums ir apmēram 10 dienas.
Mitohondriju skaita palielināšanās notiek, pieaugot un sadalot iepriekšējās mitohondrijas. Šo pieņēmumu vispirms izteica Altmans (1893), kurš aprakstīja mitohondrijas ar terminu “bioblasti”. In vivo var novērot sadalījumu, garo mitohondriju sadalīšanos īsākajos ar sašaurinājumu, kas atgādina baktēriju dalīšanas bināro metodi.
Pētot mitohondriju izturēšanos audu kultūras dzīvās šūnās, tika konstatēts reāls mitohondriju skaita pieaugums dalot. Šūnu cikla laikā mitohondriji izaug līdz vairākiem mikroniem, un pēc tam sadrumstalojas, sadalās mazākos ķermeņos.
Mitohondriji var saplūst viens ar otru un vairoties pēc principa: mitohondriji no mitohondrijiem.
Mitohondriju pašražošana.
Divu membrānu organellām ir pilnīga automātiskās reproducēšanas sistēma. Mitohondrijās un plastidās ir DNS, uz kurām tiek sintezētas informācijas, pārneses un ribosomu RNS un ribosomas, kas veic mitohondriju un plastidu olbaltumvielu sintēzi. Tomēr, lai arī šīs sistēmas ir autonomas, to iespējas ir ierobežotas.
Mitohondrijās esošā DNS ir cikliska molekula bez histoniem un tādējādi atgādina baktēriju hromosomas. To izmērs ir aptuveni 7 mikroni, vienā dzīvnieku ciklokā molekulā nonāk 16-19 tūkstoši nukleotīdu pāru DNS. Cilvēkos mitohondriju DNS satur 16,5 tūkstošus n.p., tas ir pilnībā atšifrēts. Tika konstatēts, ka dažādu priekšmetu mitohondrālās DNS ir ļoti viendabīgas, to atšķirība ir tikai intronu un nerakstīto vietu lielumā. Visas mitohondriju DNS ir vairākas kopijas, kas samontētas grupās, kopās. Tātad vienā žurku aknu mitohondrijos var būt no 1 līdz 50 cikliskām DNS molekulām. Kopējais mitohondriju DNS daudzums vienā šūnā ir aptuveni viens procents. Mitohondriju DNS sintēze nav saistīta ar DNS sintēzi kodolā.
Tāpat kā baktērijās, mitohondrāli DNS tiek savākti atsevišķā zonā - nukleoīdā, tā izmērs ir aptuveni 0,4 mikroni diametrā. Garās mitohondrijās var būt no 1 līdz 10 nukleoīdiem. Sadalot garo mitohondriju, no tā tiek atdalīta sekcija, kas satur nukleoīdu (līdzīgi kā baktēriju binārā dalīšana). DNS daudzums atsevišķos mitohondriju nukleoīdos var svārstīties 10 reizes atkarībā no šūnu veida.
Dažās kultūrās šūnās no 6 līdz 60% mitohondriju nav nukleoīdu, ko var izskaidrot ar faktu, ka šo organoīdu sadalīšana, visticamāk, ir saistīta ar fragmentāciju, nevis ar nukleoīdu sadalījumu.
Kā jau minēts, mitohondriji var gan sadalīties, gan saplūst viens ar otru. Kad mitohondriji saplūst viens ar otru, var notikt to iekšējo komponentu apmaiņa.
Svarīgi uzsvērt, ka mitohondriju un citoplazmas rRNS un ribosomas krasi atšķiras. Ja citoplazmā tiek atrastas 80. gadu ribosomas, tad augu šūnu mitohondriju ribosomas pieder 70. gadu ribosomām (sastāv no 30. un 50. subvienībām, satur 16 un 23 RNS, kas raksturīgas prokariotu šūnām), un mazākas ribosomas ir atrodamas dzīvnieku šūnu mitohondrijās (apmēram 50 sekundes).
Mitohondriju ribosomu RNS tiek sintezētas uz mitohondriju DNS. Mitoplazmā uz ribosomām notiek olbaltumvielu sintēze. Pretstatā sintēzei uz citoplazmas ribosomām tas apstājas antibiotikas hloramfenikola darbības rezultātā, kas nomāc olbaltumvielu sintēzi baktērijās.
Mitohondriju genomā tiek sintezētas 22 transporta RNS. Mitohondriju sintētiskās sistēmas trīskāršais kods atšķiras no tā, ko izmanto hialoplazmā. Neskatoties uz šķietami visu olbaltumvielu sintēzei nepieciešamo sastāvdaļu klātbūtni, mazās mitohondriju DNS molekulas nevar kodēt visus mitohondriju olbaltumvielas, tikai nelielu daļu no tām. Tātad DNS 15 tūkstošu n.p. var kodēt olbaltumvielas ar kopējo molekulmasu apmēram 6x105. Tajā pašā laikā mitohondriju pilnīga elpošanas ansambļa daļiņu olbaltumvielu kopējā molekulmasa sasniedz apmēram 2x10 6.
Ņemot vērā to, ka mitohondrijos papildus oksidatīvajiem fosforilēšanās proteīniem ietilpst arī trikarbonskābes cikla enzīmi, DNS un RNS sintēzes fermenti, aminoskābju aktivēšanas fermenti un citi proteīni, mēs redzam, ka, lai kodētu šos daudzos proteīnus un rRNS un tRNS, ģenētiskās informācijas daudzums īsās mitohondriju DNS molekulas acīmredzami trūkst. Atšifrējot cilvēka mitohondriju DNS nukleotīdu secību, parādījās, ka tā kodē tikai 2 ribosomālas RNS, 22 pārneses RNS un kopumā 13 dažādas polipeptīdu ķēdes.
Tagad ir pierādīts, ka lielākā daļa mitohondriju olbaltumvielu atrodas ģenētiskā kontrolē no šūnas kodola un tiek sintezētas ārpus mitohondrijiem. Lielākā daļa mitohondriju olbaltumvielu tiek sintezētas citosolā esošajās ribosomās. Šīm olbaltumvielām ir īpašas signālu sekvences, kuras atpazīst receptori uz mitohondriju ārējās membrānas. Šīs olbaltumvielas var ievietot tajās (skat. Analoģiju ar peroksisomu membrānu), un pēc tam pāriet uz iekšējo membrānu. Šī pārnešana notiek ārējās un iekšējās membrānas saskares vietās, kur šāds transports ir atzīmēts. Lielākā daļa mitohondriju lipīdu tiek sintezēti arī citoplazmā.
Tas viss norāda uz mitohondriju endosimbiotisko izcelsmi, ka mitohondriji ir organismi, piemēram, baktērijas, kas atrodas simbiozē ar eikariotu šūnu.
Chondriom.
Visu mitohondriju kopumu vienā šūnā sauc par hondriomu. Atkarībā no šūnu veida tas var būt atšķirīgs. Daudzās šūnās hondriomas sastāv no izkaisītām daudzām mitohondrijām, kas vienmērīgi atrodas visā citoplazmā vai lokalizētas grupās intensīvu ATP izdevumu vietās. Abos šajos gadījumos mitohondriji darbojas atsevišķi, viņu kopīgais darbs, iespējams, tiek koordinēts ar dažiem citoplazmas signāliem. Pastāv pilnīgi atšķirīgs hondriomas tips, kad mazu vienreizēju izkliedētu mitohondriju vietā šūnā atrodas viena milzu sazarota mitohondrija.
Šādas mitohondrijas ir atrodamas vienšūnu zaļajās aļģēs (piemēram, Chlorella). Tie veido sarežģītu mitohondriju tīklu vai mitohondriju retikulumu (Reticulum miyochondriale). Saskaņā ar ķīmij osmozes teoriju, šādas milzu sazarotas mitohondriju struktūras parādīšanās bioloģiskā nozīme, kuru apvieno tās ārējās un iekšējās membrānas, ir tāda, ka ATP sintēze var notikt jebkurā šādas sazarotās mitohondriju iekšējās membrānas virsmas punktā, kas nonāks jebkur citoplazmā, kur Tas ir vajadzīgs.
Milzīgu sazarotu mitohondriju gadījumā uz tā iekšējās membrānas var uzkrāties potenciāls, kas ir pietiekams ATP sintēzes sākšanai. No šīm pozīcijām mitohondriju retikulums it kā ir elektrības vadītājs, kabelis, kas savieno šādas sistēmas attālākos punktus. Mitohondriju retikulums bija ļoti noderīgs ne tikai mazām mobilajām šūnām, piemēram, chlorella, bet arī lielākām struktūrvienībām, piemēram, miofibrillēm skeleta muskuļos.
Ir zināms, ka skeleta muskuļi sastāv no muskuļu šķiedru masas, simplātiem, kas satur daudzus kodolus. Šādu muskuļu šķiedru garums sasniedz 40 mikronus ar biezumu 0,1 mikroni - tā ir gigantiska struktūra, kurā ir ļoti daudz miofibrilu, kas visi saraujas vienlaicīgi, sinhroni. Kontrakcijai liels daudzums ATP tiek piegādāts uz katru kontrakcijas vienību miofibrilu, ko mitohondriji nodrošina z-disku līmenī. Skeleta muskuļu gareniskajos ultra plānākajos sektoros elektronu mikroskops rāda neskaitāmas noapaļotas mazas mitohondriju sekcijas, kas atrodas sarkomeru tuvumā. Muskuļu mitohondriji nav mazas bumbiņas vai stieņi, bet zirnekļveidīgo struktūras, kuru procesi sazarojas un iziet lielos attālumos, dažreiz visā muskuļu šķiedras diametrā.
Tajā pašā laikā mitohondriju sazarojumi ieskauj katru muskuļu šķiedru miofibrilu, apgādājot tos ar muskuļu kontrakcijai nepieciešamo ATP. Tāpēc z-diska plaknē mitohondriji pārstāv tipisku mitohondriju retikulumu. Šādu mitohondriju retikuluma slāni vai grīdu atkārto divas reizes katram sarkomēram, un visai muskuļu šķiedrai ir tūkstošiem mitohondriju retikulāra šķērseniski novietotu “grīdas” slāņu. Starp miofibrilu “stāviem” ir šķiedru mitohondriji, kas savieno šos mitohondriju slāņus. Tādējādi tika izveidots mitohondriju retikulāra trīsdimensiju attēls, kas iet caur visu muskuļu šķiedru tilpumu.
Tālāk tika noskaidrots, ka starp mitohondriju retikulārajiem zariem un pavedienveida gareniskajām mitohondrijām ir speciāli intermitohondri savienojumi vai kontakti (MMC). Tos veido cieši blakus esošās mitohondriju saskares mitohondriju membrānas, starpmembrānu telpai un membrānām šajā zonā ir palielināts elektronu blīvums. Izmantojot šos īpašos veidojumus, kaimiņu mitohondriju un mitohondriju retikulumu funkcionālā savienība notiek vienā, sadarbīgā enerģijas sistēmā. Visas muskuļu šķiedras miofibrillas tiek samazinātas sinhroni visā garumā, tāpēc ATP piegādei jebkurai šīs sarežģītās mašīnas daļai vajadzētu notikt arī sinhroni, un tas var notikt tikai tad, ja viens ar otru ir savienots milzīgs skaits sazarotu mitohondriju vadītāju. izmantojot kontaktus.
Kardiomiocītos, sirds muskuļa šūnās bija iespējams tas, ka mitohondriju enerģētiskajā asociācijā ir iesaistīti starpmitohrālie kontakti (MMK).
Sirds muskuļa šūnu hondriomas neveido sazarotas struktūras, bet tās pārstāv daudzas mazas iegarenas mitohondrijas, kas atrodas bez īpaša pasūtījuma starp miofibrilām. Tomēr visi kaimiņu mitohondriji savienojas ar tāda paša veida mitohondriju kontaktiem kā skeleta muskuļos, tikai to skaits ir ļoti liels: vidēji katrā mitohondrijā ir 2–3 MMC, kas mitohondrijus saista vienā ķēdē, kur katrai saitei ir ķēde (Streptio mitochondriale) ir atsevišķa mitohondriji.
Izrādījās, ka starpmitohondriju kontakti (MMC) kā obligāta sirds šūnu struktūra tika atrasti visu mugurkaulnieku gan kambara, gan priekškambaru kardiomiocītos: zīdītājiem, putniem, rāpuļiem, abiniekiem un kaulainām zivīm. Turklāt MMC tika atrasti (bet mazākā skaitā) dažu kukaiņu un gliemju sirds šūnās.
MMC daudzums kardiomiocītos atšķiras atkarībā no sirds funkcionālās slodzes. MMC skaits palielinās, palielinoties dzīvnieku fiziskajai aktivitātei, un, tieši otrādi, kad samazinās sirds muskuļa slodze, krasi samazinās MMC skaits.