Lipīdu biosintēze bioķīmija. Bioķīmijas lipīdi
Elpošanas procesu starpprodukti kalpo kā oglekļa skeletu avots lipīdu sintēzei - taukiem līdzīgām vielām, kas ir visu dzīvo šūnu sastāvdaļa un spēlē nozīmīgu lomu dzīvības procesos. Lipīdi darbojas gan kā uzglabāšanas vielas, gan kā membrānu sastāvdaļas, kas ieskauj citoplazmu un visas šūnu organellus.
Membrānas lipīdi atšķiras no parastajiem taukiem ar to, ka viena no trim taukskābēm to molekulā ir aizstāta ar fosforilētu serīnu vai holīnu.
Tauki atrodas visās augu šūnās, un, tā kā tauki ūdenī nešķīst, tie augos nevar pārvietoties. Tāpēc tauku biosintēzei jānotiek visos augu orgānos un audos no izšķīdušām vielām, kas nonāk šajos orgānos. Šādas šķīstošās vielas ir ogļhidrāti, kas nokļūst sēklās no asimilācijas *. Labākais objekts tauku biosintēzes pētīšanai ir eļļas augu sēklu augļi, eļļas augu sēklu attīstības sākumā sēklu galvenie komponenti ir ūdens, olbaltumvielas, neolbaltumvielas slāpekļa savienojumi un nešķīstošie cukuri. Nogatavināšanas laikā, no vienas puses, notiek olbaltumvielu sintēze no slāpekļa savienojumiem, kas nav proteīni, un, no otras puses, ogļhidrāti pārvēršas taukos.
Mēs pievērsīsimies ogļhidrātu pārvēršanai taukos. Sāksim ar kaut ko vienkāršu. No tauku sastāva. Tauki sastāv no glicerīna un taukskābēm. Acīmredzot tauku biosintēzes laikā ir jāveidojas šiem komponentiem - glicerīnam un taukskābēm, kas ir tauku sastāvdaļa. Tauku biosintēzes laikā tika atklāts, ka taukskābes tiek kombinētas nevis ar saistīto glicerīnu, bet gan ar tā fosforilēto * - glicerīna-3 fosfātu. Glicerīna-3 fosfāta veidošanās izejmateriāls ir 3-fosfogliceraldehīds un fosfodioksiacetons, kas ir fotosintēzes un ogļhidrātu anaerobās sadalīšanās starpprodukti.
Fosfodioksiacetona reducēšanu par glicerīna-3 fosfātu katalizē enzīms glicerīna fosfāta dehidrogenāze, kura aktīvā grupa ir nikotīnamīda adenīna dinukleotīds. Taukskābju sintēze notiek sarežģītākos veidos. Mēs esam redzējuši, ka lielākajai daļai augu taukskābju ir pāra oglekļa atomu skaits, C16 vai C18. Šis fakts jau sen ir piesaistījis daudzu pētnieku uzmanību. Vairākkārt izskanējis viedoklis, ka taukskābes var veidoties etiķskābes vai acetaldehīda brīvas kondensācijas rezultātā, t.i. no savienojumiem ar diviem oglekļa atomiem C2. Mūsu laika darbos konstatēts, ka taukskābju biosintēzē piedalās nevis brīvā etiķskābe, bet ar koenzīmu A saistīts acetilkoenzīms A. Šobrīd modē ir taukskābju sintēzes shēmu attēlot šādi. Taukskābju sintēzes izejas savienojums ir acetilkoenzīms A, kas ir galvenais ogļhidrātu anaerobās sadalīšanās produkts. Koenzīms A var piedalīties dažādu taukskābju sintēzē. Pirmais * no šiem procesiem ir skābju aktivizēšana ATP iedarbībā. Pirmajā posmā no etiķskābes veidojas acetilkoenzīms A, iedarbojoties uz enzīmu acetilkoenzīms A * un patērējot enerģiju ATP un pēc tam *, t.i. notiek acetil-CoA karboksilēšana un 3 oglekļa savienojuma veidošanās. Turpmākajos posmos notiek acetilkoenzīma A molekulas kondensācija.
Taukskābju sintēze notiek, saistot acetilkoenzīma molekulu A. Šis ir faktiskās taukskābju sintēzes pirmais posms.
Vispārējo tauku veidošanās ceļu no ogļhidrātiem var attēlot kā diagrammu:
glicerīna-3 fosfāts
Ogļhidrāti
Acetilkoenzīms A → taukskābju → tauki
Kā jau zināms, tauki var pārvietoties no viena auga audiem uz otru un tie tiek sintezēti tieši uzkrāšanās vietās. Rodas jautājums: kādās šūnas daļās, kādās šūnu struktūrās tās tiek sintezētas? Augu audos tauku biosintēze gandrīz pilnībā lokalizējas mitohondrijās un sferosomās. Tauku sintēzes ātrums šūnās ir cieši saistīts ar oksidatīvo procesu intensitāti, kas ir galvenie enerģijas avoti. Citiem vārdiem sakot, tauku biosintēze ir cieši saistīta ar elpošanu.
Tauku sadalīšanās visintensīvāk notiek eļļas augu sēklu dīgšanas laikā. Eļļas sēklās ir maz ogļhidrātu, un galvenās rezerves vielas tajās ir tauki. Tauki atšķiras no ogļhidrātiem un olbaltumvielām ne tikai ar to, ka to oksidēšanās rezultātā izdalās ievērojami vairāk enerģijas, bet arī ar to, ka tauku oksidēšanās rezultātā izdalās palielināts ūdens daudzums. Ja, oksidējot 1 g olbaltumvielu, rodas 0,41 g ūdens, oksidējoties no 1 g ogļhidrātu, rodas 0,55 g, tad oksidējot 1 g tauku, rodas 1,07 g ūdens. Tam ir liela nozīme jaunattīstības embrijam, īpaši, ja sēklas dīgst sausos apstākļos.
Darbos, kas saistīti ar tauku sadalīšanās izpēti, ir pierādīts, ka dīgstošās sēklās līdz ar tauku zudumu uzkrājas ogļhidrāti. Kādos veidos ogļhidrātus var sintezēt no taukiem? Kopumā šo procesu var attēlot šādi. Tauki tiek sadalīti glicerīnā un taukskābēs ar lipāzes palīdzību, piedaloties ūdenim. Glicerīns tiek fosforilēts, pēc tam oksidēts un pārveidots par 3-fosfogliceraldehīdu. 3-fosfogliceraldehīds izomerizējas, veidojot fosfodioksiacetonu. Tālāk * un 3-fosfogliceraldehīda un fosfodioksiacetona ietekmē tiek sintezēts fruktozes-1,6 difosfāts. Izveidotais fruktozes-1,6 difosfāts, kā mēs jau zinām, tiek pārveidots par ļoti dažādiem ogļhidrātiem, kas kalpo augu šūnu un audu veidošanai.
Kāds ir taukskābju transformācijas ceļš, kas tiek atdalītas lipāzes iedarbības laikā uz taukiem? Pirmajā posmā taukskābes reakcijas ar koenzīmu A un ATP rezultātā tiek aktivizētas un veidojas acetilkoenzīms A.
R CH 2 CH 2 COOH + HS-CoA + ATP → RCH 2 CH 2 C- S – CoA
Aktivētā taukskābe, acetilkoenzīms A, ir reaktīvāka nekā brīvā taukskābe. Turpmākajās reakcijās visa taukskābes oglekļa ķēde tiek sadalīta acetilkoenzīma A divu oglekļa fragmentos. Vispārējo tauku sadalīšanas shēmu vienkāršotā veidā var attēlot šādi.
Secinājums par tauku sadalīšanās sintēzi. Gan taukskābju sadalīšanās, gan sintēzes laikā galvenā loma ir acetilkoenzīmam A. Taukskābju sadalīšanās rezultātā izveidojies acetilkoenzīms A tālāk var piedzīvot dažādas pārvērtības. Galvenais tā transformācijas ceļš ir pilnīga oksidēšanās caur trikarbonskābes ciklu līdz CO 2 un H 2 O, atbrīvojot lielu enerģijas daudzumu. Daļu no acetilkoenzīma A var izmantot ogļhidrātu sintēzei. Šādas acetilkoenzīma A transformācijas var notikt eļļas augu sēklu dīgšanas laikā, kad taukskābju aminoskābju sadalīšanās rezultātā veidojas ievērojams daudzums etiķskābes. Ogļhidrātu biosintēzes laikā no acetilkoenzīma A OH, t.i. acetilkoenzīms A ir iekļauts tā sauktajā glioksilāta ciklā jeb glioksskābes ciklā. Glioksilāta ciklā izocitrskābe tiek sadalīta dzintarskābē un glioksskābē. Dzintarskābe var piedalīties trikarbonskābes cikla reakcijā un caur * veidot ābolskābi un pēc tam skābeņetiķskābi. Glioksinskābe nonāk CO savienojumos ar otru acetilkoenzīma A molekulu, un rezultātā veidojas arī ābolskābe. Turpmākajās reakcijās ābolskābe tiek pārveidota par skābeņ-etiķskābi - fosfoenolpiruvskābi - fosfoglicerīnskābi un pat ogļhidrātos. Tādējādi sadalīšanās laikā radušos acetāta molekulas skābju enerģija tiek pārvērsta ogļhidrātos. Kāda ir glioksilāta cikla bioloģiskā loma? Šī cikla reakcijās tiek sintezēta glioksilskābe, kas kalpo kā izejas savienojums aminoskābes glicīna veidošanai. Galvenā loma ir glioksilāta cikla pastāvēšanas dēļ, taukskābju sadalīšanās laikā radušās acetāta molekulas tiek pārvērstas ogļhidrātos. Tādējādi ogļhidrātus var veidot ne tikai no glicerīna, bet arī no taukskābēm. Galīgo fotosintēzes asimilācijas produktu, ogļhidrātu, saharozes un cietes sintēze fotosintēzes šūnā tiek veikta atsevišķi: saharoze tiek sintezēta citoplazmā, ciete veidojas hloroplastos.
Secinājums. Cukurus var fermentatīvi pārveidot no viena uz otru, parasti ar ATP piedalīšanos. Ogļhidrāti tiek pārvērsti taukos, izmantojot sarežģītu bioķīmisko reakciju ķēdi. Ogļhidrātus var sintezēt no tauku sadalīšanās produktiem. Ogļhidrātus var sintezēt gan no glicerīna, gan no taukskābēm.
Aterosklerozes profilakse, kā arī slimības ārstēšana ir tieši saistīta ar lipīdu struktūru līmeņa kontroli organismā. Īpaša uzmanība tiek pievērsta holesterīnam (CS), kura molekula ir lipofils spirts. No šejienes nāk ikdienā neparastais, bet ķīmiski pareizais vielas nosaukums – holesterīns. Tā ir ķermeņa neizmantoto lipīdu oksidēšana ar brīvajiem radikāļiem, kas ir pirmais posms aterosklerozes plāksnīšu veidošanās secībā. Savukārt lipīdu struktūru savienojumi ar olbaltumvielām veido bioloģiskus kompleksus, kas spēj attīrīt asinsvadus. Tie ir augsta blīvuma lipoproteīni – ABL. Tādējādi lipīdu sintēze un biosintēze ir svarīga attiecībā uz vispārējo cilvēka veselību. Process tieši ietekmē holesterīna līmeni organismā.
Kas ietilpst lipīdu klasē?
Kategorijā ietilpst tauki un līdzīgas vielas. Molekulārā līmenī lipīds veidojas no diviem pamatelementiem: spirta un taukskābes. Ir atļautas arī papildu sastāvdaļas. Šādas struktūras pieder komplekso lipīdu klasei. No aterosklerozes profilakses viedokļa vislielāko interesi izraisa šādi šīs klases pārstāvji:
- Taukskābju spirti, proti, holesterīns.
- Triglicerīdi.
Taukskābes (FA), jo īpaši polinepiesātinātās - Omega-3, ir pelnījušas zināmu uzmanību. Viela palīdz samazināt holesterīna līmeni. Tomēr cilvēka ķermenis tos nesintezē.
Lipīdu biosintēzes vispārīgais princips
Citoplazmā sākas FA un to atvasinājumu veidošanās. Šūnā turpinās arī otrā biosintēzes daļa – molekulārās ķēdes pagarināšana, bet “ražošanas cehs nobīdās” mitohondriju iekšienē. Katrā posmā savienojums tiek bagātināts ar diviem C atomiem, kas atgādina beta oksidācijas procesu, tikai otrādi.
Sīkāk, sintēze, piemēram, palmitīnskābes, notiek tieši citoplazmā. Savukārt mitohondriji izmanto gatavu “pusfabrikātu”, lai ražotu pilnīgas taukskābes, kas sastāv no 18 vai vairāk oglekļa atomiem. Mitohondriji nespēj veikt visu biosintēzi neatkarīgi no “A” līdz “Z”. Iemesls ir banāls - “zems kvalifikācijas līmenis”. Atgriežoties pie tehniskās terminoloģijas, mitohondrijiem ir ļoti zema spēja iekļaut marķētās etiķskābes garās ķēdes lipīdu struktūrās.
Gudrs triks jeb kā metabolīts pārvar mitohondriju barjeru
Gluži pretēji, FA ekstramitohondriālajai biosintēzei nav kopīgas krustošanās ar to oksidācijas procesu. Tās mehānismam ir nepieciešami trīs komponenti:
- acetil-CoA ir primārais metabolīts;
- CO2 – šeit nav komentāru, labi zināma viela;
- bikarbonāta joni – HCO3-.
Metabolīts ir ēkas pamats. Acetil-CoA sākotnēji veidojas mitohondrijās. Tā sintēze ir oksidatīvās dekarboksilēšanas procesa sekas. Savienojums nevar tieši iekļūt citoplazmā mitohondriju membrānas necaurlaidības dēļ. Ir iespējams iekļūt, izmantojot risinājuma manevru:
- Mitohondriju metabolīts veido citrātu, mijiedarbojoties ar oksaloacetātu.
- Sintezētam citrātam mitohondriju membrāna ir caurspīdīga. Tāpēc tā molekulas viegli iekļūst citoplazmā.
- Tad notiek apgrieztā transformācija. Tik tikko šķērsojis membrānu, citrāts tiek sadalīts tā sākotnējās sastāvdaļās - acetil-CoA un oksaloacetātā.
Tādējādi metabolīts tiek pārnests no mitohondrijiem. Citoplazmā savienojuma tieša ražošana nenotiek. Ir iespējama alternatīva acetil-CoA pārnešana, piedaloties karnitīnam. Tomēr sintēzes procesā LC ir sava veida "bruņu vilciens, kas stāv uz apmales". Šis kanāls tiek izmantots daudz retāk.
Pēdējais biosintēzes posms
Nokļūstot citoplazmā, metabolīts ir gatavs FA prekursora - malonil-CoA - ražošanai. Šim nolūkam acetil-CoA nepieciešams oglekļa dioksīds. Procesa katalizators ir enzīms acetil-CoA karboksilāze. Biosintēze ir sadalīta divos periodos:
- Biotīna enzīma karboksilēšana. Rodas CO2 un ATP klātbūtnē.
- Karboksilgrupas pārnešana uz metabolītu.
Iegūtais malonil-CoA pēc tam tiek ātri pārveidots par FA. Process notiek, piedaloties noteiktai enzīmu sistēmai. Faktiski tas ir savstarpēji saistītu enzīmu komplekss. To sauc par taukskābju sintetāzi, tajā ir 6 dažādi enzīmi un savienojošais elements – acilpārneses proteīns (pilda CoA līdzīgu lomu).
Izprotot lipīdu biosintēzi vispārīgā līmenī, ir pienācis laiks pāriet pie konkrētiem piemēriem.
Triglicerīdu biosintēze
Procesa pamatelementi ir glicerīns un FA. Sākumā veidojas starpprodukts - glicerīna-3-fosfāts. Tas ir raksturīgi biosintēzes procesiem, kas notiek nierēs un zarnu sienās. Orgānu šūnām ir raksturīga glicerīna kināzes enzīma hiperaktivitāte, ko nevar teikt par muskuļiem un taukaudiem. Šeit viela veidojas, izmantojot glikolīzi - glikozes oksidēšanu.
Holesterīna biosintēze
Fermentatīvais holesterīna veidošanās process ir diezgan sarežģīts “vairāku caureju kombinācija”, kurā ir vairāk nekā 35 enzīmu reakcijas. Acīmredzami, ka tādu pārvērtību apjomu nespēj aptvert pat Ostaps Benders. Tāpēc ir vieglāk apsvērt holesterīna biosintēzes pamatposmus:
- Mevalonskābes sagatavošana. Sastopams eikariotos - dzīvo organismu domēnā. Nepieciešamas trīs aktīvā acetāta molekulas.
- Skvalēna veidošanās. Prekursors ir iepriekš ražota mevalonskābe. Sākotnēji savienojums tiek pārveidots par aktīvu izoprenoīdu, no kura 6 molekulām veidojas skvalēns.
- Holesterīna sintēze. Process tiek veikts ar skvalēna ciklizāciju. Tiek sintezēts unikāls prekursors - lanosterols, kura pāreja uz holesterīnu joprojām tiek pētīta.
Biosintēzi sākotnēji ierosina acetoacetil-CoA veidošanās. Pēc tam struktūra tiek kondensēta ar 3. aktīvā acetāta molekulu. Iegūtais atvasinājums tiek pakļauts reducēšanas reakcijai, kas izraisa mevalonāta veidošanos.
Lipīdu biosintēze
Triacilglicerīni ir viskompaktākais enerģijas uzkrāšanas veids organismā. To sintēze galvenokārt tiek veikta no ogļhidrātiem, kas organismā nonāk pārmērīgi un netiek izmantoti glikogēna krājumu papildināšanai.
Lipīdi var veidoties arī no aminoskābju oglekļa skeleta. Veicina taukskābju un pēc tam triacilglicerīnu un liekā pārtikas veidošanos.
Taukskābju biosintēze
Oksidācijas laikā taukskābes pārvēršas par acetil-CoA. Pārmērīgu ogļhidrātu uzņemšanu ar uzturu pavada arī glikozes sadalīšanās piruvātā, kas pēc tam tiek pārveidots par acetil-CoA. Šī pēdējā reakcija, ko katalizē piruvāta dehidrogenāze, ir neatgriezeniska. Acetil-CoA tiek transportēts no mitohondriju matricas uz citozolu kā daļa no citrāta (15. attēls).
Mitohondriju matricas citozols
15. attēls. Acetil-CoA pārneses shēma un reducētā NADPH veidošanās taukskābju sintēzes laikā.
Stereoķīmiski visu taukskābju sintēzes procesu var attēlot šādi:
Acetil-CoA + 7 Malonil-CoA + 14 NADPH∙ + 7H +
Palmitīnskābe (C 16:0) + 7 CO 2 + 14 NADP + 8 NSCoA + 6 H 2 O,
šajā gadījumā no acetil-CoA veidojas 7 malonil-CoA molekulas:
7 Acetil-CoA + 7 CO 2 + 7 ATP 7 Malonil-CoA + 7 ADP + 7 H 3 PO 4 + 7 H +
Malonil-CoA veidošanās ir ļoti svarīga reakcija taukskābju sintēzē. Malonil-CoA veidojas acetil-CoA karboksilēšanas reakcijā, piedaloties acetil-CoA karboksilāzei, kas satur biotīnu kā protezēšanas grupu. Šis enzīms neietilpst taukskābju sintāzes multienzīmu kompleksā. Acetīta karboksilāze ir polimērs (molekulmasa no 4 līdz 810 6 Da), kas sastāv no protomēriem ar molekulmasu 230 kDa. Tas ir daudzfunkcionāls allosterisks proteīns, kas satur saistītu biotīnu, biotīna karboksilāzi, transkarboksilāzi un alosterisko centru, kura aktīvā forma ir polimērs, un 230 kDa protomēri ir neaktīvi. Tāpēc malonil-CoA veidošanās aktivitāti nosaka attiecība starp šīm divām formām:
Neaktīvie protomēri aktīvs polimērs
Palmitoil-CoA, biosintēzes galaprodukts, novirza attiecību pret neaktīvo formu, un citrāts, būdams allosteriskais aktivators, novirza šo attiecību pret aktīvo polimēru.
16. attēls. Malonil-CoA sintēzes mehānisms
Karboksilēšanas reakcijas pirmajā posmā tiek aktivizēts bikarbonāts un veidojas N-karboksibiotīns. Otrajā posmā notiek acetil-CoA karbonilgrupas N-karboksibiotīna nukleofils uzbrukums un transkarboksilēšanas reakcijā veidojas malonil-CoA (16. att.).
Taukskābju sintēze zīdītājiem ir saistīta ar multienzīmu kompleksu, ko sauc taukskābju sintāze.Šo kompleksu attēlo divi identiski daudzfunkcionāli polipeptīdi. Katram polipeptīdam ir trīs domēni, kas atrodas noteiktā secībā (att.). Pirmais domēns ir atbildīgs par acetil-CoA un malonil-CoA saistīšanu un šo divu vielu savienošanu. Šajā domēnā ietilpst enzīmi acetiltransferāze, maloniltransferāze un acetilmalonilu saistošs enzīms, ko sauc par β-ketoacilsintazi. Otrais domēns, galvenokārt ir atbildīgs par pirmajā domēnā iegūtā starpprodukta reducēšanu un satur acilpārneses proteīnu (ACP), -ketoacilreduktāzi un dehidrāzi un enoil-ACP reduktāzi. IN trešais domēns ir enzīms tioesterāze, kas atbrīvo iegūto palmitīnskābi, kas sastāv no 16 oglekļa atomiem.
Rīsi. 17. Palmitāta sintāzes kompleksa uzbūve. Cipari norāda domēnus.
Taukskābju sintēzes mehānisms
Taukskābju sintēzes pirmajā posmā acetiltransferāzes serīna atlikumam pievieno acetil-CoA (att....). Līdzīgā reakcijā starp malonil-CoA un maloniltransferāzes serīna atlikumu veidojas starpprodukts. Pēc tam acetilgrupa no acetiltransferāzes tiek pārnesta uz acilpārneses proteīna (ATP) SH grupu. Nākamajā posmā acetila atlikums tiek pārnests uz -ketoacilsintāzes cisteīna (kondensējošā enzīma) SH grupu. Acilpārneses proteīna brīvā SH grupa uzbrūk maloniltransferāzei un saistās ar malonila atlikumu. Tad notiek malonila un acetila atlikumu kondensācija, piedaloties -ketoacilsintazei ar karbonilgrupas atdalīšanu no malonila. Reakcijas rezultātā veidojas -ketoacils, kas saistīts ar ĀKK.
Rīsi. 3-ketoacilACP sintēzes reakcijas palmitāta sintāzes kompleksā
Pēc tam otrā domēna fermenti piedalās β-ketoacil-ACP starpprodukta reducēšanas un dehidratācijas reakcijās, kuru rezultātā veidojas (butiril-ACP) acil-ACP.
Acetoacetil-ACP (-ketoacil-ACP)
-ketoacil-ACP reduktāze
-Hidroksibutiril-APB
-hidroksiacil-ACP dehidrāze
Enoil-ĀKK reduktāze
Butirils-APB
Pēc 7 reakcijas cikliem
H2O palmitoiltioesterāze
Pēc tam butirilgrupa tiek pārnesta no ĀKK uz -ketoacilsintāzes cis-SH atlikumu. Tālāka pagarināšana ar diviem oglekļa atomiem notiek, maloniltransferāzes serīna atlikumam pievienojot malonil-CoA, pēc tam atkārtojas kondensācijas un reducēšanas reakcijas. Viss cikls tiek atkārtots 7 reizes un beidzas ar palmitoil-ACP veidošanos. Trešajā domēnā palmitoilesterāze hidrolizē tioestera saiti par palmitoil-ACP, un tiek atbrīvota brīvā palmitīnskābe, kas atstāj palmitāta sintāzes kompleksu.
Taukskābju biosintēzes regulēšana
Taukskābju sintēzes kontrole un regulēšana zināmā mērā ir līdzīga glikolīzes, citrāta cikla un taukskābju β-oksidācijas reakciju regulēšanai. Galvenais metabolīts, kas iesaistīts taukskābju biosintēzes regulēšanā, ir acetil-CoA, kas nāk no mitohondriju matricas kā daļa no citrāta. Malonil-CoA molekula, kas veidojas no acetil-CoA, inhibē karnitīna aciltransferāzi I, un taukskābju β-oksidācija kļūst neiespējama. No otras puses, citrāts ir acetil-CoA karboksilāzes allosteriskais aktivators, un palmitoil-CoA, steatoril-CoA un arahidonil-CoA ir galvenie šī enzīma inhibitori.
LIPĪDI.BIOLS.LOMA.KLASIFIKĀCIJA.
Lipīdi ir liela bioloģiskas izcelsmes vielu grupa, kas labi šķīst organiskos šķīdinātājos, piemēram, metanolā, acetonā, hloroformā un benzolā. Lipīdi ir vissvarīgākais visu uzturvielu enerģijas avots. Šūnu membrānu veidošanā piedalās vairāki lipīdi.Daži lipīdi veic īpašas funkcijas organismā. Steroīdi, eikozanoīdi un daži fosfolipīdu metabolīti veic signalizācijas funkcijas. Tie kalpo kā hormoni, neirotransmiteri un sekundārie transportētāji. Lipīdi ir sadalīti pārziepjojamos un nepārziepjojamos. Pārziepjojamie lipīdi.
Pārziepjojamos lipīdos ietilpst trīs vielu grupas: esteri, fosfolipīdi un glikolipīdi. Estera grupā ietilpst neitrālie tauki, vaski un sterīnu esteri.Fosfolipīdu grupā ietilpst fosfatīdskābes, fosfatīdi un sfingolipīdi.Glikolipīdu grupā ietilpst cerebrozīdi un gangliozīdi).
Nepārziepjojamo lipīdu grupā ietilpst piesātinātie ogļūdeņraži un karotinoīdi, kā arī spirti. Pirmkārt, tie ir spirti ar garu alifātisku ķēdi, cikliskie sterīni (holesterīns) un steroīdi (estradiols, testosterons utt.). Vissvarīgāko lipīdu grupu veido taukskābes. Šajā grupā ietilpst arī eikozanoīdi, kurus var uzskatīt par taukskābju atvasinājumiem.
Lipīdu sagremošana un lipīdu gremošanas produktu uzsūkšanās.
Mutes dobumā tauki neizmainās, jo Siekalas nesatur fermentus, kas šķeļ taukus. Lai gan pieauguša cilvēka kuņģī nenotiek būtiska pārtikas tauku sagremošana, kuņģī joprojām tiek novērota pārtikas šūnu membrānu lipoproteīnu kompleksu daļēja iznīcināšana, kas padara taukus pieejamākus turpmākai aizkuņģa dziedzera sulas lipāzes iedarbībai uz tiem. Tauku, kas veido pārtiku, sadalīšanās notiek cilvēkiem un zīdītājiem galvenokārt tievās zarnas augšdaļās, kur ir ļoti labvēlīgi apstākļi tauku emulgācijai. Pēc tam, kad chyme nokļūst divpadsmitpirkstu zarnā, šeit, pirmkārt, tiek neitralizēta kuņģa sulas sālsskābe. Taukskābes ar īsu oglekļa ķēdi un glicerīnu, kas labi šķīst ūdenī, brīvi uzsūcas zarnās un nokļūst portāla vēnas asinīs un no turienes aknās, apejot jebkādas pārvērtības zarnu sieniņās. Garā leņķa taukskābes ķēde ir grūtāk absorbējama. Ar žults, žults sāļu, fosfolipīdu un holesterīna palīdzību. Micellas, kas brīvi uzsūcas zarnās.
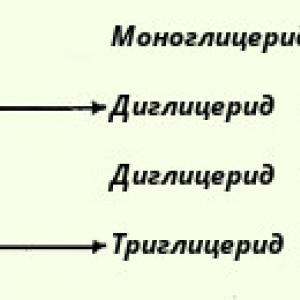
3. Triacilglicerīdu hidrolīze. Tauku resintēze.
Triacilglicerīdi ir visizplatītākie lipīdi dabā. Tos parasti iedala taukos un eļļās. Triacilglicerīnu hidrolīze rada glicerīnu un taukskābes. Pilnīga triglicerīdu hidrolīze notiek pa posmiem: pirmkārt, 1. un 3. saite tiek strauji hidrolizēta, un tad 2-monoglicerīda hidrolīze notiek lēni.. (hidrolīze). Tauku resintēze zarnu sieniņās. Zarnu sieniņās tiek sintezēti tauki, kas lielā mērā ir raksturīgi konkrētai dzīvnieku sugai un pēc būtības atšķiras no uztura taukiem. Triglicerīdu resintēzes mehānisms zarnu sieniņu šūnās kopumā izpaužas šādi: sākotnēji to aktīvā forma acil-CoA veidojas no taukskābēm, pēc tam notiek monoglicerīdu acilēšana, veidojoties pirmajiem diglicerīdiem. un tad triglicerīdi: 
4. Žultsskābes.struktūra, bioloģiskā loma.
Žultsskābes veidojas no holesterīna aknās. Šie 24 oglekļa steroīdu savienojumi ir holānskābes atvasinājumi, kuriem ir viena līdz trīs α-hidroksilgrupas un sānu ķēde ar 5 oglekļa atomiem ar karboksilgrupu ķēdes galā. Holskābe ir vissvarīgākā skābe cilvēka organismā.  Žultsskābes nodrošina holesterīna šķīdību žultī un veicina lipīdu gremošanu.
Žultsskābes nodrošina holesterīna šķīdību žultī un veicina lipīdu gremošanu.
Lipīdu un to sastāvdaļu biosintēze.
Paši lipīdi un daži to strukturālie komponenti cilvēka organismā nonāk galvenokārt ar pārtiku. Ja nav pietiekama lipīdu piegāde no ārpuses, organisms spēj daļēji novērst lipīdu komponentu deficītu to biosintēzes ceļā. Tādējādi dažas piesātinātās skābes organismā var sintezēt fermentatīvi. Zemāk redzamā diagramma atspoguļo palmitīnskābes veidošanās procesa no etiķskābes kopsavilkumu:
CH3COOH + 7HOOC - CH2 - COOH + 28 [H]
C15H31COOH + 7CO2 + 14H2O
Šis process tiek veikts, izmantojot koenzīmu A, kas pārvērš skābes tioesteros un aktivizē to dalību nukleofilās aizvietošanas reakcijās:
Dažas nepiesātinātās skābes (piemēram, oleīns un palmitoleīns) cilvēka organismā var sintezēt, dehidrogenējot piesātinātās skābes. Linolskābes un linolēnskābes cilvēka organismā netiek sintezētas un nāk tikai no ārpuses. Galvenais šo skābju avots ir augu pārtika. Linolskābe kalpo kā avots arahidonskābes biosintēzei. Tā ir viena no svarīgākajām skābēm, kas veido fosfolipīdus.Triacilglicerīnus un fosfatīdskābes sintezē uz glicero-3-fosfāta bāzes, kas veidojas no glicerīna, pāresterificējoties ar ATP. No kopējā holesterīna daudzuma organismā tikai 20% nāk ar pārtiku. Galvenais holesterīna daudzums organismā tiek sintezēts, piedaloties koenzīmam acetil-CoA.
Lipīdus prokariotu šūnā attēlo dažāda rakstura ķīmiskie savienojumi (triglicerīdi, fosfolipīdi, glikolipīdi, vaski), kas veic dažādas funkcijas. Tie ir daļa no šūnu membrānām, ir pigmentu sistēmu un elektronu transporta sastāvdaļas un darbojas kā uzglabāšanas vielas. Lipīdu biosintēzes sākumprodukti ir taukskābes, spirti, ogļhidrāti un fosfāti. Lipīdu biosintēzes ceļi ir sarežģīti un prasa ievērojamu enerģijas daudzumu, piedaloties daudziem fermentiem. Triglicerīdi un fosfolipīdi ir vissvarīgākie šūnu funkcionēšanai.
Taukskābju ar pāra oglekļa atomu skaitu biosintēze notiek, secīgi pievienojot malonil-CoA divu oglekļa atlikumu acetil-CoA molekulai. Tādējādi palmitīnskābes biosintēzes laikā 1 acetil-CoA molekula tiek kondensēta ar 7 malonil-CoA molekulām:
Acetil-CoA + 7 malonil-CoA + 14 NAD(P)H 2
CH 3 (CH 2) 14 COOH +7 CO 2 + 8CoA + 14NAD(P) + +6H 2 O
Svarīgu lomu taukskābju biosintēzes reakcijās spēlē acilpārneses proteīns (ATP), acilgrupu nesējs. Secīgs divu oglekļa atlieku pieaugums, izmantojot virkni starpproduktu, izraisa C16-C18 savienojumu veidošanos. Prokariotu šūnās lipīdu komponenti var būt nepiesātinātās taukskābes, kas satur vienu dubultsaiti. Divkāršās saites veidošanās aerobos mikroorganismos notiek ar skābekļa un specifiska enzīma desaturāzes piedalīšanos. Piemēram, palmitoleīnskābe veidojas no palmitil-CoA:
Palmitil-CoA + ½ O 2 + NAD(P)H 2 palmitooleil-CoA + H 2 O + NAD(P) +
Anaerobos mikroorganismos dubultsaites veidošanās notiek taukskābju molekulas biosintēzes agrīnā stadijā dehidratācijas reakcijas rezultātā.
Sākuma substrāts fosfolipīdu sintēzei ir fosfodioksiacetons, glikolītiskā cikla starpprodukts. Tā samazināšana noved pie 3-fosfoglicerīna veidošanās, kas, savienojoties ar divām taukskābju atlikumiem, rada fosfatidskābi. Serīna, inozīna, etanolamīna un holīna pievienošana tās fosfātu grupai izraisa fosfatidilserīna, fosfatidilinozīta, fosfatidilholīna un fosfatidiletanolamīna sintēzi.