책:
물의 밀도- 이것은 수중 생물의 이동 조건과 다른 깊이에서의 압력을 결정하는 요소입니다. 증류수의 경우 밀도는 4°C에서 1g/cm3입니다. 용해된 염을 함유한 자연수의 밀도는 최대 1.35g/cm3까지 높을 수 있습니다. 압력은 수심에 따라 평균 10m마다 약 1 · 10 5 Pa(1 atm)씩 증가합니다.
수역의 급격한 압력 구배 때문에 수생 생물은 일반적으로 육상 생물보다 훨씬 더 유황성입니다. 다른 깊이에서 흔히 볼 수 있는 일부 종은 몇 기압에서 수백 기압까지 견딜 수 있습니다. 예를 들어, Elpidia 속의 해삼, Priapulus caudatus 벌레는 해안 지역에서 초심연까지 서식합니다. 예를 들어 섬모류, 슬리퍼, suvoy, 수영 딱정벌레 등과 같은 민물 거주자도 실험에서 최대 6 · 10 7 Pa(600 atm)를 견딜 수 있습니다.
그러나 바다와 바다의 많은 주민들은 상대적으로 협착증을 앓고 있으며 특정 깊이에 국한되어 있습니다. Stenobathism은 얕은 물과 깊은 물 종의 특징입니다. 연안 지역에만 고리형 모래벌레 Arenicola와 바다 접시(Patella)가 서식합니다. 예를 들어 낚시꾼 그룹의 많은 물고기, 두족류, 갑각류, 포고노포어, 불가사리 등은 최소한 4 · 10 7 - 5 · 10 7 Pa(400-500 atm)의 압력에서 깊은 곳에서만 발견됩니다.
물의 밀도는 물에 의존할 수 있는 능력을 제공하며, 이는 골격 형태에 특히 중요합니다. 환경의 밀도는 물에서 치솟는 조건으로 작용하며 많은 수중 생물이 이 특정한 생활 방식에 적응합니다. 매달린 수중 유기체는 수생 유기체의 특별한 생태 그룹으로 결합됩니다. 플랑크톤 ( "플랑크톤"- 급등).
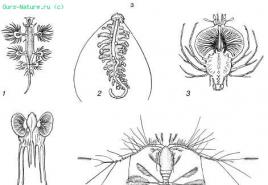
쌀. 39. 플랑크톤 유기체에서 신체의 상대 표면 증가(S. A. Zernov, 1949에 따름):
A - 막대 모양:
1 - 규조류 Synedra;
2 - 시아노박테리움 아파니조메논;
3 - Peridineal alga Amphisolenia;
4 - 유글레나 아쿠스;
5 - 두족류 Doratopsis vermicularis;
6 - 요각류 세텔라;
7 - 포셀라나 유충(Decapoda)
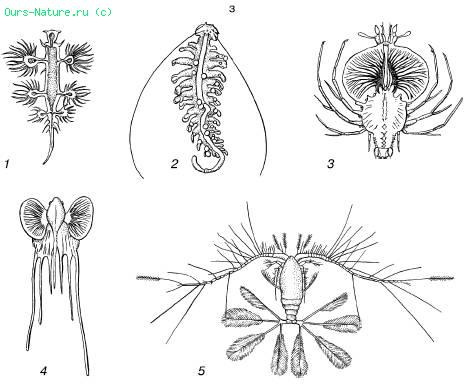
B - 절단된 형태:
1 - 연체 동물 Glaucus atlanticus;
2 - 웜 Tomopetris euchaeta;
3 - 가재 Palinurus의 유충;
4 - 아귀 Lophius의 물고기의 유충;
5 - 요각류 Calocalanus pavo
플랑크톤에는 단세포 및 군체 조류, 원생동물, 해파리, 사이펀포어, 빗해파리, 익룡류 및 용골 연체동물, 다양한 작은 갑각류, 저서 동물의 유충, 생선 알 및 치어 및 기타 많은 것들이 포함되어 있습니다(그림 39). 플랑크톤 유기체는 부력을 증가시키고 바닥으로 가라앉는 것을 방지하는 유사한 적응을 많이 합니다. 이러한 적응에는 다음이 포함됩니다. 1) 크기 감소, 평평함, 신장, 물에 대한 마찰을 증가시키는 수많은 파생물 또는 강모의 발달로 인한 신체의 상대 표면의 일반적인 증가; 2) 골격 감소로 인한 밀도 감소, 체내 지방 축적, 기포 등 규조류에서는 예비 물질이 무거운 전분의 형태가 아니라 지방 방울의 형태로 침착됩니다. Nightlight Noctiluca는 세포에 풍부한 가스 액포와 지방 방울이 있어서 세포질이 핵 주위에서만 합쳐지는 가닥처럼 보입니다. Siphonophores, 많은 해파리, 플랑크톤 복족류 등에도 공기 챔버가 있습니다.
해초 (식물성 플랑크톤)대부분의 플랑크톤 동물은 능동적으로 수영할 수 있지만 제한된 한계 내에서 수동적으로 물에 떠 있습니다. 플랑크톤 유기체는 조류를 극복할 수 없으며 조류에 의해 장거리로 운반됩니다. 많은 종류 동물성 플랭크톤그러나 활발한 움직임과 몸의 부력 조절로 인해 수십 미터와 수백 미터의 수주에서 수직 이동이 가능합니다. 특별한 유형의 플랑크톤은 생태 그룹입니다. 뉴스톤 ( "Nein"- 수영) - 공기 환경과의 경계에서 물 표면 필름의 주민.
물의 밀도와 점도는 능동적으로 수영하는 능력에 큰 영향을 미칩니다. 빠르게 헤엄칠 수 있고 조류의 힘을 극복할 수 있는 동물들이 하나의 생태 그룹으로 결합됩니다. 유영 동물 ( "Nektos"- 부동). 넥톤의 대표자는 물고기, 오징어, 돌고래입니다. 물기둥에서의 빠른 움직임은 유선형의 몸매와 고도로 발달된 근육이 있어야만 가능합니다. 어뢰 모양은 체계적인 소속 및 수중 움직임 방법에 관계없이 모든 우수한 수영 선수에서 개발됩니다.
산소 모드.산소가 함유 된 물에서 그 함량은 1 리터당 10ml를 초과하지 않으며 이는 대기보다 21 배 낮습니다. 따라서 수생 생물의 호흡 조건은 상당히 복잡합니다. 산소는 주로 조류의 광합성 활동과 공기로부터의 확산을 통해 물에 들어갑니다. 따라서 수주의 상층은 일반적으로 하층보다이 가스가 풍부합니다. 물의 온도와 염분이 증가함에 따라 물의 산소 농도가 감소합니다. 동물과 박테리아가 많이 서식하는 층에서는 소비 증가로 인해 급격한 O 2 결핍이 발생할 수 있습니다. 예를 들어, 세계 해양에서 생명이 풍부한 50-1000m 깊이는 폭기가 급격히 악화되는 특징이 있습니다. 이는 식물성 플랑크톤이 서식하는 지표수보다 7-10배 낮습니다. 수역의 바닥 근처의 조건은 혐기성에 가까울 수 있습니다.
수중 거주자 중에는 물이 거의 완전히 없을 때까지 물의 산소 함량이 크게 변동하는 것을 견딜 수 있는 많은 종이 있습니다. (유로옥시비온트 - "옥시"- 산소, "biont"- 거주자). 여기에는 예를 들어 담수 oligochaetes Tubifex tubifex 및 복족류 Viviparus viviparus가 포함됩니다. 물고기 중에서 잉어, 텐치, 붕어 등은 매우 약한 산소 포화도를 견딜 수 있습니다. 그러나 여러 유형의 스테녹시바이오트 - 그들은 산소로 충분히 높은 포화도에서만 존재할 수 있습니다 (무지개 송어, 갈색 송어, 미노, 섬모 벌레 Planaria alpina, 하루살이의 유충, 돌파리 등). 많은 종은 산소 부족으로 비활성 상태로 떨어질 수 있습니다. 무산소증 - 따라서 불리한 기간을 경험합니다.
수생 생물의 호흡은 신체의 표면이나 아가미, 폐, 기관과 같은 특수 기관을 통해 수행됩니다. 이 경우 외피는 추가 호흡기 역할을 할 수 있습니다. 예를 들어 미꾸라지 물고기는 평균적으로 피부를 통해 최대 63%의 산소를 소비합니다. 신체의 외피를 통해 가스 교환이 발생하면 매우 얇습니다. 또한 표면적이 넓어져 호흡이 쉬워집니다. 이것은 다양한 파생물의 형성, 편평화, 길어짐 및 신체 크기의 일반적인 감소에 의해 종의 진화 중에 달성됩니다. 산소가 부족한 일부 종은 호흡 표면의 크기를 적극적으로 변경합니다. Tubifex tubifex 벌레는 몸을 강하게 늘입니다. 히드라와 말미잘 - 촉수; 극피 동물 - 보행 다리. 많은 앉아있는 동물과 앉아있는 동물은 물의 방향 흐름을 만들거나 물의 혼합에 기여하는 진동 운동을 통해 주변의 물을 재생합니다. 이 목적을 위해, 이매패류는 맨틀 구멍의 벽을 감싸는 섬모입니다. 갑각류 - 복부 또는 흉부 다리의 작업. 거머리, 벨모기(혈충)의 유충, 많은 oligochaetes가 땅에서 튀어나와 몸을 흔들립니다.
일부 종에서는 물과 공기 호흡의 조합이 발견됩니다. 이들은 폐 호흡 물고기, siphonophores disophants, 많은 폐 연체 동물, 갑각류 Gammarus lacustris 등입니다. 2 차 동물은 일반적으로 대기 유형의 호흡을 에너지 적으로 더 유익한 것으로 유지하므로 공기 환경과의 접촉이 필요합니다. 예를 들어 기각류, 고래류, 물 딱정벌레, 모기 유충 및 기타.
물의 산소 부족은 때때로 치명적인 현상으로 이어집니다. 자모라, 많은 수중 생물의 죽음을 동반합니다. 겨울잠종종 저수지 표면에 얼음이 형성되고 공기와의 접촉이 종료되어 발생합니다. 여름- 결과적으로 수온의 상승과 산소 용해도의 감소.
겨울에 물고기와 많은 무척추 동물의 빈번한 죽음은 예를 들어 Ob 강 유역의 하부에서 전형적입니다. 그 물은 서쪽 시베리아 저지의 늪지대에서 흘러 내리는 물은 용존 산소가 극도로 부족합니다. 때로는 바다에서 죽음이 발생합니다.
산소 부족 외에도 바닥에서 유기 물질이 분해되어 형성되는 메탄, 황화수소, CO 2 등의 유독 가스 농도 증가로 인해 서리가 발생할 수 있습니다. 저수지.
소금 모드.수생 생물의 물 균형을 유지하는 데에는 고유한 특성이 있습니다. 육상 동물과 식물의 경우 결핍 상태에서 몸에 물을 제공하는 것이 가장 중요하다면 수생 생물의 경우 환경에서 과잉으로 몸에 일정량의 물을 유지하는 것도 똑같이 중요합니다. 세포의 과도한 양의 물은 삼투압의 변화와 가장 중요한 생명 기능의 붕괴를 초래합니다.
대부분의 수중 생물 포이킬로스모틱: 그들의 몸의 삼투압은 주변 물의 염도에 따라 달라집니다. 따라서 수생 생물이 염분 균형을 유지하는 주요 방법은 염분이 부적절한 서식지를 피하는 것입니다. 민물 형태는 바다에 존재할 수 없으며 바다 형태는 담수화를 견딜 수 없습니다. 물의 염도가 변하면 동물들은 유리한 환경을 찾아 움직입니다. 예를 들어, 폭우 후 바다 표층의 담수화 중에 방사성 유충, 바다 갑각류 Calanus 등은 100m 깊이로 내려갑니다. 척추 동물, 고등 가재, 곤충 및 물에 사는 유충은 항상성 물의 염분 농도에 관계없이 체내에서 일정한 삼투압을 유지하는 종.
담수 종에서 바디 주스는 주변 물과 관련하여 고장성입니다. 과도한 물이 몸에 들어가거나 제거되는 것을 방지하지 않으면 과도한 물을 줄 위험이 있습니다. 원생 동물에서 이것은 배설 시스템을 통해 물을 제거함으로써 다세포 유기체에서 배설 액포의 작업에 의해 달성됩니다. 일부 섬모는 2-2.5분마다 신체의 부피와 동일한 양의 물을 방출합니다. 세포는 과도한 물을 "펌핑"하기 위해 많은 에너지를 소비합니다. 염분이 증가함에 따라 액포의 작용이 느려집니다. 따라서 염분이 2.5% o인 Paramecium 신발에서 액포는 9초 간격으로 5% o - 18초, 7.5% o - 25초 간격으로 맥동합니다. 17.5% o의 염 농도에서 세포와 환경 사이의 삼투압 차가 사라지기 때문에 액포가 작동을 멈춥니다.
물이 수생 유기체의 체액과 관련하여 고장성인 경우 삼투 손실의 결과로 탈수 위험이 있습니다. 탈수에 대한 보호는 수생 생물체의 염분 농도를 증가시켜 달성됩니다. 탈수는 포유류, 물고기, 고등 가재, 수생 곤충 및 그 유충과 같은 동종 삼투성 유기체의 불투수성 덮개에 의해 방지됩니다.
많은 poikilosmotic 종은 염분 증가와 함께 체내 수분 부족으로 인해 비활성화 상태로 전환됩니다. 이것은 로티퍼, 편모류, 섬모류, 일부 갑각류, 흑해 다모류 Nereis divesicolor 등 해수 웅덩이와 연안 지역에 사는 종의 전형입니다. 염분 아나비시스- 물의 다양한 염도 조건에서 불리한 기간 동안 생존하는 수단.
진실로 유리염활성 상태의 민물과 바닷물 모두에서 살 수있는 수생 거주자 중에는 그렇게 많은 종이 있지 않습니다. 이들은 주로 하천 하구, 하구 및 기타 기수 수역에 서식하는 종입니다.
온도 체제수역은 육지보다 더 안정적입니다. 이는 물의 물리적 특성, 주로 높은 비열 용량 때문이며, 이로 인해 상당한 양의 열을 받거나 방출해도 온도가 너무 급격하게 변화하지 않습니다. 약 2263.8 J / g이 소비되는 저수지 표면에서 물의 증발은 하층의 과열을 방지하고 융해열 (333.48 J / g)이 방출되는 얼음 형성은 그들의 속도를 늦춥니다. 냉각.
해양 상층의 연간 온도 변동의 진폭은 대륙 수역 - 30-35 ° C에서 10-15 ° C 이하입니다. 깊은 수층은 일정한 온도가 특징입니다. 적도 수역에서 표층의 평균 연간 온도는 + (26-27) ° C이고 극지방에서는 약 0 ° C 이하입니다. 뜨거운 육지 온천에서 수온은 +100 ° C에 도달 할 수 있으며 해저의 고압 수중 간헐천에서는 +380 ° C의 온도가 기록됩니다.
따라서 수역에는 상당히 다양한 온도 조건이 있습니다. 계절적 온도 변동이 있는 물의 상층과 열 체제가 일정한 하층 사이에는 온도 점프 또는 수온약층이 있습니다. 수온약층은 외부와 심해의 온도차가 더 강한 따뜻한 바다에서 더 두드러집니다.
수생 생물 사이의 보다 안정적인 수온 체계로 인해 stenotherm은 육지 개체군보다 훨씬 더 광범위하게 퍼져 있습니다. Eurythermal 종은 주로 얕은 대륙 수역과 일별 및 계절별 온도 변동이 큰 고위도 및 온대 위도의 연안에서 발견됩니다.
라이트 모드.물에는 공기보다 빛이 훨씬 적습니다. 저수지 표면에 떨어지는 광선의 일부는 공기 중으로 반사됩니다. 태양의 위치가 낮을수록 반사가 강해지기 때문에 육지보다 수면 아래에서 낮이 짧습니다. 예를 들어, 마데이라 섬 근처의 여름날 수심 30m - 5시간, 수심 40m에서는 단 15분. 깊이에 따라 빛의 양이 급격히 감소하는 것은 물에 의한 흡수와 관련이 있습니다. 파장이 다른 광선은 같은 방식으로 흡수되지 않습니다. 빨간색은 이미 표면 가까이에서 사라지고 청록색은 훨씬 더 깊숙이 침투합니다. 바다에 깊어지는 황혼은 처음에는 녹색이고, 그 다음에는 파란색, 파란색, 청자색이 되며 마침내 끝없는 어둠에 자리를 내줍니다. 따라서 파장이 다른 빛을 포착하는 데 특화되어 있는 녹조류, 갈조류, 홍조류는 깊이로 서로를 대체합니다.
동물의 색깔도 같은 방식으로 깊이에 따라 변합니다. 연안 및 연안 지역의 주민들은 가장 생생하고 다양한 색을 띠고 있습니다. 동굴 생물과 같은 많은 심부 생물체에는 색소가 없습니다. 황혼 영역에서는 붉은 색이 널리 퍼져 있으며, 이는 이 깊이의 청자색 빛을 보완합니다. 보색 광선은 신체에 가장 완전히 흡수됩니다. 이것은 청자색 광선의 붉은 색이 시각적으로 검은 색으로 인식되기 때문에 동물이 적으로부터 숨을 수 있습니다. 붉은 색은 농어, 붉은 산호, 다양한 갑각류 등과 같은 황혼 지대의 동물에 일반적입니다.
수면 근처에 사는 일부 종에서 눈은 광선을 굴절시키는 능력이 다른 두 부분으로 나뉩니다. 눈의 절반은 공중에서 보고 다른 절반은 물 속에서 봅니다. 이러한 "네 눈"은 바다 개 Dialommus fuscus의 열대 종 중 하나인 딱정벌레 빙빙 도는 미국 물고기 Anableps tetraphthalmus의 전형입니다. 썰물 때 이 물고기는 오목한 곳에 앉아 머리의 일부를 물 밖으로 내보냅니다(그림 26 참조).
빛의 흡수가 강할수록 물의 투명도는 낮아지며, 이는 물에 부유하는 입자의 양에 따라 다릅니다.
투명도는 직경이 약 20cm인 특별히 낮아진 흰색 디스크(Secchi 디스크)가 여전히 보이는 극도의 깊이가 특징입니다. 가장 투명한 물은 Sargasso Sea에 있습니다 : 디스크는 66.5m 깊이까지 볼 수 있으며 태평양에서 Secchi 디스크는 최대 59m, 인도양에서는 최대 50m, 얕은 바다에서는 최대 50m까지 볼 수 있습니다. 5-15m 강의 투명도는 평균 1-1 , 5m이며 가장 진흙 투성이의 강, 예를 들어 중앙 아시아 Amu Darya 및 Syrdarya에서는 불과 몇 센티미터입니다. 따라서 광합성 구역의 경계는 수역에 따라 크게 다릅니다. 가장 깨끗한 물에서 환희의광합성 구역 또는 광합성 구역이 200m 이하, 황혼 또는 난시,이 영역은 최대 1000-1500m 깊이를 차지하며 더 깊습니다. 무극성영역, 햇빛이 전혀 침투하지 않습니다.
수역 상층부의 빛의 양은 지역의 위도와 계절에 따라 크게 달라집니다. 긴 극의 밤은 북극과 남극 분지의 광합성에 사용할 수 있는 시간을 심각하게 제한하고 얼음으로 덮인 겨울에는 빛이 모든 얼어붙은 수역에 접근하기 어렵습니다.
바다의 어두운 깊이에서 유기체는 생물이 방출하는 빛을 시각 정보의 소스로 사용합니다. 살아있는 유기체의 빛을 호출합니다. 생물발광.발광 종은 원생동물에서 어류에 이르기까지 거의 모든 종류의 수생 동물뿐만 아니라 박테리아, 하등 식물 및 균류에서 발견됩니다. 생물발광은 분명히 진화의 여러 단계에서 여러 그룹에서 여러 번 나타났습니다.
생물발광의 화학은 이제 상당히 잘 이해되었습니다. 빛을 생성하는 데 사용되는 반응은 다양합니다. 그러나 모든 경우에 이것은 복잡한 유기 화합물의 산화입니다. (루시페린)단백질 촉매를 사용하여 (루시페라제).다른 유기체의 루시페린과 루시페라제는 다른 구조를 가지고 있습니다. 반응하는 동안 여기된 루시페린 분자의 과잉 에너지가 빛 양자의 형태로 방출됩니다. 살아있는 유기체는 일반적으로 외부 환경의 자극에 반응하여 펄스로 빛을 방출합니다.
발광은 종의 생활에서 특별한 생태학적 역할을 하지 않을 수 있지만, 예를 들어 박테리아나 하등 식물에서와 같이 세포의 중요한 활동의 부산물일 수 있습니다. 그것은 충분히 발달 된 신경계와 시력 기관을 가진 동물에서만 생태 학적 중요성을받습니다. 많은 종에서 발광 기관은 방사선을 증폭시키는 반사경 및 렌즈 시스템으로 매우 복잡한 구조를 얻습니다(그림 40). 빛을 생성할 수 없는 많은 물고기와 두족류는 이러한 동물의 특수 기관에서 번식하는 공생 박테리아를 사용합니다.
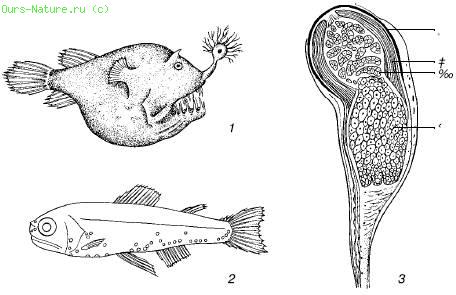
쌀. 40. 수생 동물의 발광 기관(S.A.Zernov, 1949에 따름):
1 - 이빨이 있는 입 위에 손전등이 있는 심해 낚시꾼
2 - 이 물고기의 발광 기관 분포. 미스토피과;
3 - 물고기 Argyropelecus affinis의 발광 기관:
a - 안료, b - 반사경, c - 발광체, d - 렌즈
생물 발광은 주로 동물의 삶에서 중요한 신호입니다. 빛 신호는 무리에서 방향을 제시하여 이성을 유인하거나 희생자를 유인하거나 위장 또는 주의를 산만하게 할 수 있습니다. 섬광은 포식자로부터 눈을 멀게 하거나 방향 감각을 잃게 하는 방어 수단이 될 수 있습니다. 예를 들어, 적으로부터 도주하는 심해 오징어는 빛나는 분비물의 구름을 방출하는 반면 밝은 물에 사는 종은 이러한 목적으로 어두운 액체를 사용합니다. 일부 저서 벌레 - 갯지렁이 - 발광 기관은 생식 산물의 성숙기에 발달하며 암컷보다 밝게 빛나고 눈은 수컷에서 더 잘 발달합니다. 아귀목의 육식성 심해 어류에서 등지느러미의 첫 번째 광선은 위턱으로 이동하고 끝에 벌레 같은 "미끼"를 운반하는 유연한 "막대"로 바뀝니다. 빛나는 박테리아. 동맥으로 가는 혈류를 조절하고 결과적으로 박테리아에 산소를 공급함으로써 물고기는 임의로 "미끼"를 빛나게 하여 벌레의 움직임을 모방하고 먹이를 유인할 수 있습니다.