Как называют внутренние структуры митохондрий. Строение и функции митохондрий и пластид
Митохондрии
В клетках животных тканей митохондрии были обнаружены в 1882 г., а у растений - только в 1904 г. (в пыльниках кувшинки). Биологические функции удалось установить после выделения и очистки фракции методом фракционного центрифугирования. В их составе находится 70% белка и около 30% липидов, небольшое количество РНК и ДНК, витамины А, B 6 , В 12 , К, Е, фолиевая и пантотеновая кислоты, рибофлавин, различные ферменты. Митохондрии имеют двойную мембрану, Наружная изолирует органеллу от цитоплазмы, а внутренняя образует выросты кристы. Все пространство между мембранами заполнено матриксом (рис. 13).
Основная функция митохондрий - участие в клеточном дыхании. Роль митохондрий в дыхании была установлена в 1950-1951 годах. На наружных мембранах концентрируется сложная ферментная система цикла Кребса. При окислении субстратов дыхания освобождается энергия, которая тотчас же в процессе окислительного фосфорилирования, происходящего в кристах, аккумулируется в образующихся молекулах АДФ и главным образом АТФ. Энергия, запасенная в макроэргических соединениях, используется в дальнейшем для удовлетворения всех потребностей клетки.
Образование митохондрий в клетке происходит непрерывно из микротелец, чаще их возникновение связывают с дифференцировкой мембранных структур клетки. Они в клетке могут восстанавливаться путем их деления и почкования. Митохондрии не долговечны, продолжительность их жизни 5-10 дней.
Митохондрии – «силовые» станции клетки. В них концентрируется энергия, которая запасается в «аккумуляторах» энергии - молекулах АТФ, а не рассеивается в клетке. Нарушение структуры митохондрии ведет к нарушению процесса дыхания и в итоге к патологии организма.
Аппарат Гольджи. Аппарат Гольджи (синоним - диктиосомы) представляет собой стопки из 3-12 уплощенных, замкнутых, окруженных двойной мембраной дисков, называемых цистернами, от краев которых отшнуровываются многочисленные пузырьки (300-500). Ширина цистерн 6-90 А, толщина мембран - 60-70 А.
Аппарат Гольджи является центром синтеза, накопления и выделения полисахаридов, в частности целлюлозы, участвует в распределении и внутриклеточном транспорте белков, а также в образовании вакуолей и лизосом. В растительной клетке удалось проследить участие аппарата Гольджи в возникновении срединной пластинки и росте клеточной пекто-целлюлозной оболочки.
Аппарат Гольджи более всего развит в период активной жизни клетки. При ее старении он постепенно атрофируется, а затем исчезает.
Лизосомы. Лизосомы - довольно мелкие (около 0.5 мк в диаметре) округлые тельца. Они покрыты белково-липоидной мембраной. Содержимое лизосом многочисленные гидролитические ферменты, которые осуществляют функцию внутриклеточного переваривания (лизирования) макромолекул белка, нуклеиновых кислот, полисахаридов. Их основная функция переваривание отдельных участков протопласта клетки (автофагия - самопожирание). Этот процесс протекает за счет фагоцитоза или пиноцитоза. Биологическая роль этого процесса двоякая. Во-первых, защитная, поскольку при временном недостатке запасных продуктов клетка поддерживает жизнь за счет конституционных белков и др. веществ, а во-вторых происходит освобождение от избыточных или изношенных органелл (пластид, митохондрий и др.) Оболочка лизосомы препятствует выходу ферментов в цитоплазму, в противном случае она бы вся переваривалась этими ферментами.
В умершей клетке лизосомы разрушаются, ферменты оказываются в клетке и все ее содержимое переваривается. Остается только пекто-целлюлозная оболочка.
Лизосомы являются продуктами деятельности аппарата Гольджи, оторвавшимися от него пузырьками, в которых этот органоид аккумулировал переваривающие ферменты.
Сферосомы - округлые белково-липоидные тельца 0.3-0.4 мкм. По всей вероятности являются производными аппарата Гольджи или эндоплазматического ретикулума. По своей форме и величине напоминают лизосомы. Поскольку сферосомы содержат кислую фосфатазу, то они, вероятно, имеют отношение к лизосомам. Некоторые авторы считают, что сферосомы и лизосомы эквивалентны друг другу, но, скорее всего только по происхождению и форме. Есть предположение об их участии в синтезе жиров (А.Фрей-Висслинг).
Рибосомы - очень мелкие органоиды, диаметр их около 250А, По форме они почти шаровидные. Часть их прикреплена к наружным мембранам эндоплазматического ретикулума, часть их находится в свободном состоянии в цитоплазме. В клетке может содержаться до 5 млн рибосом. Рибосомы есть в хлоропластах и митохондриях, где они синтезируют часть белков, из которых построены эти органоиды, и ферменты, функционирующие в них.
Основная функция - синтез специфических белков согласно информации, поступающей из ядра. Их состав: белок и рибосомная рибонуклеиновая кислота (РНК) в равных соотношениях. Их структура малая и большая субъединицы, сформированные из рибонуклеотида.
Микротрубочки. Микротрубочки - своеобразные производные эндоплазматического ретикулума. Обнаружены во многих клетках. Само их название говорит об их форме - одна или две, расположенные параллельно, трубочки с полостью внутри. Внешний диаметр в пределах 250А. Стенки микротрубочек построены из белковых молекул. Из микротрубочек во время деления клетки образуются нити веретена.
Ядро
Ядро было обнаружено в растительной клетке Р. Броуном в 1831 году. Оно располагается в центре клетки или около клеточной оболочки, но со всех сторон окружено цитоплазмой. В большинстве случаев в клетке находится одно ядро, по несколько ядер находится в клетках некоторых водорослей, а также грибов. У зеленых водорослей неклеточной структуры насчитывается сотни ядер. Многоядерные клетки нечленистых млечников. Отсутствуют ядра в клетках бактерий и сине-зеленых водорослей.
Форма ядра чаще всего близка к форме шара или эллипса. Зависит от формы, возраста и функции клетки. В меристематической клетке ядро крупное, округлой формы и занимает 3/4 объема клетки. В паренхимных клетках эпидермы, имеющих крупную центральную вакуоль, ядро имеет чечевицеобразную форму и отодвинуто вместе с цитоплазмой к периферии клетки. Это признак специализированной, но уже стареющей клетки. Клетка, лишенная ядра, способна жить лишь короткое время. Безъядерные клетки ситовидных трубок живые клетки, но живут они недолго. Во всех других случаях безъядерные клетки являются мертвыми.
Ядро имеет двойную оболочку, через поры в которой содержимое
ядра (нуклеоплазма) может сообщаться с содержимым цитоплазмы. Мембраны оболочки ядра снабжены рибосомами и сообщаются с мембранами эндоплазматического ретикулума клетки. В нуклеоплазме располагается одно или два ядрышка и хромозомы. Нуклеоплазма представляет собой коллоидную систему золя, напоминающую по консистенции загустевшую желатину. В ядре, по данным отечественных биохимиков (Збарский И.Б. и др.), содержится четыре фракции белков: простых белков - глобулинов 20%, дезоксирибонуклеопротеидов - 70%, кислых белков - 6% и остаточных белков 4%. Они локализуются в следующих ядерных структурах: ДНК-протеиды (щелочные белки) - в хромозомах, РНК-протеиды (кислые белки) - в ядрышках, частично в хромозомах (в период синтеза информационной РНК) и в ядерной мембране. Глобулины составляют основу нуклеоплазмы. Остаточные белки (природа не уточнена) образуют ядерную мембрану.
Основная масса белков ядра - сложные щелочные белки дезоксирибонуклеопротеиды, в основе которых находится ДНК.
Молекула ДНК. Молекула ДНК – полинуклеотид и состоит из нуклеотидов. В состав нуклеотида входит три компонента: молекула сахара (дезоксирибоза), молекула азотистого основания и молекулы фосфорной кислоты. Дезоксирибоза соединена с азотистым основанием гликозидной, а с фосфорной кислотой - эфирной связью. В ДНК имеется в различных комбинациях всего 4 разновидности нуклеотидов, отличающихся друг от друга азотистыми основаниям. Два из них (аденин и гуанин) относятся к пуриновым азотистым соединениям, а цитозин и тимин - к пиримидиновым. Молекулы ДНК располагаются не в одной плоскости, а состоят из двух спирализованных нитей, т.е. две параллельно расположенные цепочки, закрученные одна вокруг другой, образуют одну молекулу ДНК. Они скреплены между собой с помощью водородной связи между азотистыми основаниями, причем пуриновые основания одной цепочки присоединяют пиримидиновые основания другой (рис.14). Структура и химизм молекулы ДНК была раскрыта английским (Крик) и американским (Уотсон) учеными и обнародована в 1953 г. Этот момент принято считать началом развития молекулярной генетики. Молекулярный вес ДНК – 4-8 млн. Количество нуклеотидов (различных вариантов) до 100 тысяч. Молекула ДНК очень стабильна, ее стабильность обеспечивается тем, что на всем протяжении она имеет одинаковую толщину - 20А (8А - ширина пиримидинового основания +12А - ширина пуринового основания). Если ввести в организм радиоактивный фосфор, то метка будет обнаруживаться во всех фосфоросодержащих соединениях, кроме ДНК (Леви, Сикевиц).
Молекулы ДНК являются носителями наследственности, т.к. в их структуре закодирована информация о синтезе специфических белков, определяющих свойства организма. Изменения могут возникнуть под действием мутагенных факторов (радиоактивное излучение, сильнодействующие, химические агенты -алкалоиды, спирты и т.д.).
Молекула РНК. Молекулы рибонуклеиновой кислоты (РНК) значительно меньше молекул ДНК. Это одиночные цепочки из нуклеотидов. Существует три вида РНК: рибосомная, самая длинная, образующая многочисленные петли, информационная (матричная) и траспортная, самая короткая. Рибосомная РНК локализуется в рибосомах эндоплазматической сети и составляет 85% всей РНК клетки.
Информационная РНК по своей структуре напоминает листочек клевера. Ее количество - 5% от суммы всей РНК в клетке. Она синтезируется в ядрышках. Ее сборка происходит в хромозомах в период интерфазы. Ее основная функция - перенос информации от ДНК к рибосомам, где происходит синтез белка.
Транспортная РНК, как установлено сейчас, это целое семейство соединений, родственных по структуре и биологической функции. Каждая живая клетка по приблизительной оценке содержит 40-50 индивидуальных транспортных РНК и их общее число в природе, если учесть видовые различия, огромно. (Акад.В.Энгельгардт). Транспортными они называются потому, что их молекулы заняты транспортным обслуживанием внутриклеточного процесса синтеза белка. Соединяясь со свободными аминокислотами, они доставляют их к рибосомам в строящуюся белковую цепь. Это самые маленькие молекулы РНК, состоят в среднем из 80 нуклеотидов. Локализуются в матриксе цитоплазмы и составляют около 10% клеточной РНК
В составе РНК содержится четыре азотистых основания, но в отличие от ДНК в молекуле РНК вместо тимина находится урацил.
Структура хромозом. Хромозомы впервые были обнаружены в конце 19 века классиками цитологии Флемингом и Страсбургером (1882, 1884), а русский исследователь клетки И.Д. Чистяков их обнаружил в 1874 году.
Основной структурный элемент хромозом - ядро. Они имеют различную форму. Это либо прямые, либо изогнутые палочки, овальные тельца, шарики, размеры которых варьируют.
В зависимости от места расположения центромеры различают прямые, равноплечие и неравноплечие хромозомы. Внутренняя структура хромозом представлена на рис. 15, 16. Следует отметить, что дезоксирибонуклеопротеид является мономером хромозомы.
В хромозоме дезоксирибонуклеопротеидов 90-92%, из них 45% -ДНК и 55% - белка (гистона). В небольшом количестве в хромозоме представлена и РНК (информационная).
У хромозомы четко выражена и поперечная структура - наличие утолщенных участков - дисков, которые еще в 1909г. были названы генами. Этот термин был предложен датским ученым Иогансеном. В 1911 г. американский ученый Морган доказал, что гены являются основными наследственными единицами и распределяются они в хромозомах в линейном порядке и, поэтому хромозома имеет качественно различные участки. В 1934 г. американский ученый Пайнтер доказал прерывистость морфологического строения хромозом и наличие в хромозомах дисков, а диски - это места скопления ДНК. Это послужило началом создания хромосомных карт, на которых указывалось место (локус) расположения гена, определяющего тот или иной признак организма. Ген - это участок двойной спирали ДНК, на котором содержится информация о структуре одного белка. Это участок молекулы ДНК, определяющий синтез одной молекулы белка. ДНК непосредственного участия в синтезе белка не принимает. В ней только содержится и хранится информация о структуре белка.
Структура ДНК, состоящая из нескольких тысяч последовательно расположенных 4-х нуклеотидов, представляет собой код наследственности.
Код наследственности. Синтез белка. Первое сообщение по коду ДНК сделал американский биохимик Ниренберг в 1961 г. в Москве на международном биохимическом конгрессе. Сущность кода ДНК состоит в следующем. Каждой аминокислоте соответствует участок цепи ДНК из трех рядом расположенных нуклеотидов (триплет). Так, например участок, состоящий из Т-Т-Т (триплет из 3-х тиминсодержащих нуклеотидов) соответствует аминокислоте лизину, триплет А (аденин) - Ц (цитозин) - А (аденин)- цистеину и т.д. Допустим, что ген представлен цепочкой нуклеотидов, расположенных в следующем порядке: А-Ц-А-Т-Т-Т-А-А-Ц-Ц-А-А-Г-Г-Г. Разбив этот ряд на триплеты, мы сразу расшифруем, какие аминокислоты и в каком порядке будут располагаться в синтезируемом белке.
Число возможных сочетаний из 4-х имеющихся нуклеотидов по три равно 4×64. Исходя из этих соотношений, числа различных триплетов с избытком хватит для обеспечения информации по синтезу многочисленных белков, определяющих и структуру и функции организма. Для синтеза белка в рибосомы направляется точная копия этой информации в виде информационной РНК. В расшифровке и синтезе, кроме и-РНК, участвует большое число молекул различных транспортных рибонуклеиновых кислот (т-РНК), рибосомы и ряд ферментов. Каждая из 20 аминокислот связывается с Т-РНК - молекула с молекулой. Каждой из 20 аминокислот соответствует своя т-РНК. У т-РНК имеются химические группы, способные «узнавать» свою аминокислоту, выбирая именно ее из наличных аминокислот. Происходит это с помощью специальных ферментов. Узнав свою аминокислоту, т-РНК вступает с ней в соединение. К началу цепочки (молекулы) и-РНК присоединяется рибосома, которая, продвигаясь по и-РНК, соединяет друг с другом в полипептидную цепочку именно те аминокислоты, порядок которых зашифрован нуклеотидной последовательностью данной И-РНК. В результате образуется молекула белка, состав которого закодирован в одном из генов.
Ядрышки - неотъемлемая структурная часть ядра. Это сферические тельца. Они очень изменчивы, меняют свою форму и структуру, появляются и исчезают. Их бывает одно, два. Для каждого растения определенное число. Ядрышки исчезают, когда клетка готовится к делению, а затем появляются вновь; они, по-видимому, участвуют в синтезе рибонуклеиновых кислот. Если ядрышко разрушить сфокусированным пучком рентгеновских или ультрафиолетовых лучей, то клеточное деление подавляется.
Роль ядра в жизни клетки. Ядро служит контролирующим центром клетки- оно направляет клеточную активность и содержит носителей наследственности (гены), определяющие признаки данного организма. Роль ядра можно выявить, если с помощью микрохирургических приемов удалить его из клетки и наблюдать последствие этого. Ряд опытов, доказывающих важную роль в регуляции клеточного роста, провел Геммерлинг на одноклеточной зеленой водоросли Acetobularia. Эта морская водоросль достигает высоты 5 см, внешне напоминает гриб, имеет подобие "корней" и "ножки". Вверху заканчивается большой дисковидной "шляпкой". Клетка этой водоросли имеет одно ядро, располагающееся в базальной части клетки.
Гаммерлинг установил, что если перерезать ножку, то нижняя часть продолжает жить и полностью регенерируют шляпку после операции. Верхняя же часть, лишенная ядра, выживает в течение некоторого времени, но, в конце концов, погибает, не будучи в состоянии восстановить нижнюю часть. Следовательно, ацетобулярии ядро необходимо для метаболических реакций, лежащих в основе роста.
Ядро способствует образованию клеточной оболочки. Это можно проиллюстрировать экспериментами с водорослью Voucheria и Spyrogyra. Выпуская из перерезанных нитей в воду содержимое клеток, мы можем получить комочки цитоплазмы с одним, с несколькими ядрами и без ядер. В первых двух случаях клеточная оболочка формировалась нормально. В случае отсутствия ядра оболочка не образовывалась.
В опытах И.И.Герасимова (1890г.) со спирогирой было установлено, что клетки с двойным ядром удваивают длину и толщину хлоропласта. В безъядерных клетках продолжается процесс фотосинтеза, образуется ассимиляционный крахмал, но при этом затухает процесс его гидролиза, что объясняется отсутствием гидролитических ферментов, которые могут быть синтезированы в рибосомах лишь согласно информации ДНК ядра. Жизнь протопласта без ядра неполноценна и недолговечна. В экспериментах И.И. Герасимова безъядерные клетки спирогиры жили 42 дня и погибали. Одна из важнейших функций ядра состоит в снабжении цитоплазмы рибонуклеиновой кислотой, необходимой для синтеза белка в клетке. Удаление ядра из клетки ведет к постепенному падению содержания РНК в цитоплазме и замедлению синтеза белка в ней.
Наиболее важна роль ядра в передаче признаков от клетки клетке, от организма к организму и осуществляет это в процессе деления ядра и клетки в целом.
Клеточное деление. Размножаются клетки делением. При этом из одной клетки образуется две дочерних с тем же набором наследственного материала, заключенного в хромозомах, что и материнская клетка. В соматических клетках хромозомы представлены двумя, так называемыми гомологическими хромозомами, в которых заложены аллельные гены (носители противоположных признаков, например, белый и красный цвет лепестков гороха и т.д.), признаков двух родительских пар. В связи с этим в соматических клетках тела растения всегда удвоенный набор хромозом, обозначаемый 2п. Хромозомы обладают выраженной индивидуальностью. Количество и качество хромозом - характерный признак каждого вида. Так, в клетках земляники диплоидный набор хромозом равен 14, (2n), яблони -34, топинамбура - 102 и т.д.
Митоз (кариокинез) – деление соматических клеток был впервые описан Э. Руссовым(1872г.) и И.Д.Чистяковым (1874). Его сущность заключается в том, что из материнской клетки путем деления образуется две дочерние клетки с тем же набором хромозом.Клеточный цикл слагается из интерфазы и собственно митоза. Методом микроавторадиографии установлено, что самой длительной и сложной является интерфаза - период "покоящегося" ядра, т.к. в этот период происходит удвоение ядерного материала. Интерфаза делится на три фазы:
Q1 - пресинтетическая (ее длительность 4-6 часов);
S - синтетическая (10-20 часов);
Q2 - постсинтетическая (2-5 часов).
Во время Q1 фазы идет подготовка к редупликации ДНК. А в S-фазу происходит редупликация ДНК, клетка удваивает запас ДНК. В Q2-фазу формируются ферменты и структуры, необходимые для запуска митоза. Таким образом, в интерфазе молекулы ДНК в хромозомах расщепляются на две одинаковые нити, происходит сборка на их матрице информационных РНК. Последняя уносит информацию о структуре специфических белков в цитоплазму, а в ядре каждая из нитей ДНК достраивает недостающую половинку своей молекулы. В этом процессе удвоения (редупликация) проявляется уникальная особенность ДНК, состоящая в способности ДНК точно воспроизводить саму себя. Образовавшиеся дочерние молекулы ДНК автоматически получаются точными копиями родительской молекулы, т.к. при редупликации к каждой половинке присоединяются комплементарные (А-Т; Г-Ц; и т.д.) основания из окружающей среды.
В профазу митотического деления удвоенные хромозомы становятся заметными. В метафазе все они располагаются в экваториальной зоне, располагаясь в один ряд. Образуются нити веретена (из микротрубочек, соединяющихся друг с другом). Оболочка ядра и ядрышко исчезают. Утолщенные хромозомы расщепляются вдоль на две дочерние хромозомы. В этом заключается суть митоза. Он обеспечивает точное распределение удвоенных молекул ДНК между дочерними клетками. Тем самым обеспечивает и передачу зашифрованной в ДНК наследственной информации.
В анафазе дочерние хромозомы начинают отходить к противоположным полюсам. В центре появляются первые фрагменты клеточной оболочки (фрагмобласт).
В телофазе происходит оформление ядер в дочерних клетках. Содержимое материнской клетки (органеллы) распределяется между образующимися дочерними. Полностью формируется клеточная оболочка. На этом заканчивается цитокинез (рис.17).
Мейоз - редукционное деление был обнаружен и описан в 90-х годах прошлого столетия В.И.Беляевым. Сущность деления заключается в том, что из соматической клетки, содержащей 2п (двойной, диплоидный) набор хромозом, образуется четыре гаплоидных клетки, с"n", половинным набором хромозом. Этот тип деления является сложным и состоит из двух этапов. Первый - редукция хромозом. Удвоенные хромозомы располагаются в экваториальной зоне попарно (две параллельно расположенные гомологичные хромозомы). В этот момент может происходить коньюгация (сцепление) хромозом, кроссинговер (перекрест) и в результате - обмен участками хромозом. В результате этого часть генов отцовских хромозом переходит в состав материнских хромозом и наоборот. Внешний вид тех и других хромозом в результате этого не меняется, но их качественный состав становится иным. Отцовская и материнская наследственности перераспределяются и смешиваются.
В анафазе мейоза гомологичные хромозомы с помощью нитей веретена расходятся по полюсам, на которых после небольшого периода покоя (исчезают нити, но перегородка между новыми ядрами не формируется) начинается процесс митоза - метафаза, при которой все хромозомы располагаются в одной плоскости и происходит их продольное расщепление на дочерние хромозомы. При анафазе митоза с помощью веретена они расходятся по полюсам, где и формируется четыре ядра и в итоге - четыре гаплоидные клетки. В клетках некоторых тканей при их развитии под влиянием некоторых факторов происходит незавершенный митоз и количество хромозом в ядрах удваивается за счет того, что не расходятся по полюсам. В результате таких нарушений естественного или искусственного характера возникают организмы тетраплоиды и полиплоиды. С помощью мейоза формируются половые клетки - гаметы, а также споры, элементы полового и бесполого размножения растений (рис.18).
Амитоз - прямое деление ядра. При амитозе веретено деления не образуется и оболочка ядра не распадается, как при митозе. Раньше амитоз рассматривался как примитивная форма деления. Сейчас установлено, что он связан с деградацией организма. Представляет собой упрощенный вариант более сложного деления ядра. Амитоз встречается в клетках и тканях нуцеллуса, эндосперма, паренхиме клубней, черешков листьев и т.д.
Митохондрия - это спиральная, округлая, вытянутая или разветвленная органелла.
Впервые понятие митохондрия было предложено Бенда в 1897 г. Митохондрии можно обнаружить в живых клетках с помощью фазово-контрастной и интерференционной микроскопии в виде зерен, гранул или нитей. Это довольно подвижные структуры, которые могут перемещаться, сливаться друг с другом, делиться. При окраске специальными методами в погибших клетках при световой микроскопии митохондрии имеют вид мелких зерен (гранул), диффузно распределенных в цитоплазме или концентрирующихся в каких-то определенных ее зонах.
В результате разрушения глюкозы и жиров в присутствии кислорода в митохондриях образуется энергия, а органические вещества превращаются в воду и диоксид углерода. Именно таким образом получают основную энергию, необходимую для жизнедеятельности, животные организмы. Энергия накапливается в аденозинтрифосфате (АТФ), а точнее, в его макроэргических связях. Функция митохондрий тесно связана с окислением органических соединений и использованием освобождающейся при их распаде энергии для синтеза молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания. АТФ выполняет функцию поставщика энергии, перенося одну из своих богатых энергией концевых фосфатных групп на другую молекулу, и превращается при этом в АДФ.
Предполагают, что в эволюции митохондрии были прокариотическими микроорганизмами, которые стали симбиотами в организме древней клетки. В последующем они стали жизненно необходимы, что было связано с увеличением содержания кислорода в атмосфере Земли. С одной стороны, митохондрии удаляли избыток токсичного для клетки кислорода, а с другой - обеспечивали энергией.
Без митохондрий клетка практически не в состоянии использовать кислород как вещество, обеспечивающее поставку энергии, и может восполнять свои энергетические потребности лишь путем анаэробных процессов. Таким образом, кислород - это яд, но яд жизненно важный для клетки, причем избыток кислорода так же вреден, как и его недостаток.
Митохондрии могут изменять свою форму и перемещаться в те области клетки, где потребность в них наиболее высока. Так, в кардиомиоцитах митохондрии находятся вблизи миофибрилл, в клетках почечных канальцев вблизи базальных впячиваний и т. д. В клетке содержится до тысячи митохондрий, и их количество зависит от активности клетки.
Митохондрии имеют средние поперечные размеры 0,5…3 мкм. В зависимости от размеров выделяют мелкие, средние, крупные и гигантские митохондрии (формируют разветвленную сеть - митохондриальный ретикулум). Размеры и число митохондрий тесно связаны с активностью клетки и ее энергопотреблением. Они крайне изменчивы и в зависимости от активности клетки, содержания кислорода, гормональных влияний могут набухать, изменять число и структуру крист, варьировать в числе, форме и размерах, а также ферментативной активности.
Объемная плотность митохондрий, степень развития их внутренней поверхности и другие показатели зависят от энергетических потребностей клетки. В лимфоцитах имеется всего по несколько митохондрий, а в печеночных клетках их 2…3 тыс.
Митохондрии состоят из матрикса, внутренней мембраны, перимитохондриального пространства и наружной мембраны. Наружная митохондриальная мембрана отделяет органеллу от гиалоплазмы. Обычно она имеет ровные контуры и замкнута так, что представляет собой мембранный мешок.
Внешнюю мембрану от внутренней отделяет перимитохондриальное пространство шириной около 10…20 нм. Внутренняя митохондриальная мембрана ограничивает собственно внутреннее содержимое митохондрии - матрикс. Внутренняя мембрана образует многочисленные выпячивания внутрь митохондрий, которые имеют вид плоских гребней, или крист.
По форме кристы могут иметь вид пластинок (трабекулярные) и трубочек (мультивезикулярные на срезе), а направлены они продольно или поперечно по отношению к митохондрии.
Каждая митохондрия заполнена матриксом, который на электронных микрофотографиях выглядит плотнее, чем окружающая цитоплазма. Матрикс митохондрии однородный (гомогенный), иногда мелкозернистый, различной электронной плотности. В нем выявляют тонкие нити толщиной около 2…3 нм и гранулы размером около 15…20 нм. Нити матрикса представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы. В матриксе содержатся ферменты, одна одноцепочечная, циклическая ДНК, митохондриальные рибосомы, много ионов Са 2+ .
Автономная система белкового синтеза митохондрий представлена молекулами ДНК, свободными от гистонов. ДНК короткая, имеет форму кольца (циклическая) и содержит 37 генов. В отличие от ядерной ДНК в ней практически нет некодирующих последовательностей нуклеотидов. Особенности строения и организации сближают ДНК митохондрий с ДНК бактериальных клеток. На ДНК митохондрий происходит синтез молекул РНК разных типов: информационных, трансфертных (транспортных) и рибосомальных. Информационная РНК митохондрий не подвергается сплайсингу (вырезанию участков, не несущих информационной нагрузки). Малые размеры молекул митохондриальных ДНК не могут определить синтез всех белков митохондрий. Большинство белков митохондрий находится под генетическим контролем клеточного ядра и синтезируется в цитоплазме, так как ДНК митохондрий слабо выражена и может обеспечить образование лишь части ферментов цепи окислительного фосфорилирования. Митохондриальная ДНК кодирует не более десяти белков, которые локализованы в мембранах и представляют собой структурные белки, ответственные за правильную интеграцию отдельных функциональных белковых комплексов митохондриальных мембран. Синтезируются также белки, осуществляющие транспортные функции. Такая система белкового синтеза не обеспечивает всех функций митохондрии, поэтому автономия митохондрий ограниченная и относительная.
У млекопитающих митохондрии при оплодотворении передаются лишь через яйцеклетку, а спермий привносит в новый организм ДНК ядра.
В матриксе митохондрий образуются рибосомы, отличающиеся от рибосом цитоплазмы. Они участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром. Митохондриальные рибосомы имеют число седиментации 60 (в отличие от цитоплазматических с числом седиментации 80). Число седиментации - это скорость осаждения при центрифугировании и ультрацентрифугировании. По строению митохондриальные рибосомы близки к рибосомам прокариотических организмов, но меньшего размера и отличаются чувствительностью к определенным антибиотикам (левомицетину, тетрациклину и др.).
Внутренняя мембрана митохондрии обладает высокой степенью избирательности при транспорте веществ. К ее внутренней поверхности прикрепляются тесно прилежащие друг к другу ферменты цепи окислительного фосфорилирования, белки-переносчики электронов, транспортные системы АТФ, АДФ, пируват и др. В результате тесного расположения ферментов на внутренней мембране обеспечивается высокая сопряженность (взаимосвязанность) биохимических процессов, повышающая скорость и эффективность каталитических процессов.
При электронной микроскопии выявляют грибовидные частицы, выступающие в просвет матрикса. Они обладают АТФ-синтетичной (образует АТФ из АДФ) активностью. Транспорт электронов идет по дыхательной цепи, локализованной во внутренней мембране, которая содержит четыре крупных ферментных комплекса (цитохромы). При прохождении электронов по дыхательной цепи ионы водорода откачиваются из матрикса в перимитохондриальное пространство, что обеспечивает формирование протонного градиента (помпы). Энергия этого градиента (различия в концентрации веществ и формирование мембранного потенциала) используется для синтеза АТФ и транспорта метаболитов и неорганических ионов. Содержащиеся на внутренней мембране белки-переносчики транспортируют через нее органические фосфаты, АТФ, АДФ, аминокислоты, жирные кислоты, три — и дикарбоновые кислоты.
Наружная мембрана митохондрии более проницаема для низкомолекулярных веществ, так как в ней много гидрофильных белковых каналов. На наружной мембране располагаются специфические рецепторные комплексы, через которые белки из матрикса транспортируются в перимитохондриальное пространство.
По своему химическому составу и свойствам наружная мембрана близка к другим внутриклеточным мембранам и плазмолемме. В ней содержатся ферменты, метаболизирующие жиры, активирующие (катализирующие) превращения аминов, аминооксидаза. Если ферменты наружной мембраны сохраняют активность, то это показатель функциональной сохранности митохондрий.
В митохондриях имеются два автономных субкомпартмента. Вели перимитохондриальное пространство, или наружная камера митохондрии (внешний субкомпартмент), формируется за счет проникновения белковых комплексов гиалоплазмы, то внутренний субкомпартмент (матрикс митохондрии) частично образован за счет синтетической активности митохондриальной ДНК. Во внутреннем субкомпартменте (матриксе) содержатся ДНК, РНК и рибосомы. Он отличается высоким уровнем ионов Са 2+ в сравнении с гиалоплазмой. Во внешнем субкомпартменте накапливаются ионы водорода. Ферментативная активность внешнего и внутреннего субкомпартментов, состав белков сильно различаются. Внутренний субкомпартмент имеет более высокую электронную плотность, чем внешний.
Специфические маркеры митохондрий - ферменты цитохром-оксидаза и сукцинатдегидрогеназа, выявление которых позволяет количественно охарактеризовать энергетические процессы в митохондриях.
Основная функция митохондрий - синтез АТФ. Вначале в гиалоплазме разрушаются сахара (глюкоза) до молочной и пировиноградной кислот (пирувата) с одновременным синтезом небольшого количества АТФ. В результате гликолиза одной молекулы глюкозы используется две молекулы АТФ, а образуется четыре. Таким образом, положительный баланс составляют всего две молекулы АТФ. Эти процессы совершаются без кислорода (анаэробный гликолиз).
Все последующие этапы выработки энергии происходят в процессе аэробного окисления, который обеспечивает синтез большого количества АТФ. При этом органические вещества разрушаются до СO 2 и воды. Окисление сопровождается переносом протонов на их акцепторы. Эти реакции осуществляются с помощью ряда ферментов цикла трикарбоновых кислот, которые находятся в матриксе митохондрии.
В мембраны крист встроены системы переноса электронов и сопряженного с ним фосфорилирования АДФ (окислительное фосфорилирование). При этом происходит перенос электронов от одного белка-акцептора электронов к другому и, наконец, связывание их с кислородом, вследствие чего образуется вода. Одновременно с этим часть энергии, выделяемой при таком окислении в цепи переноса электронов, запасается в виде макроэргической связи при фосфорилировании АДФ, что приводит к образованию большого числа молекул АТФ - основного внутриклеточного энергетического эквивалента. На мембранах крист митохондрий происходит процесс окислительного фосфорилирования с помощью расположенных здесь белков цепи окисления и фермента фосфорилирования АДФ АТФ-синтетазы. В результате окислительного фосфорилирования из одной молекулы глюкозы образуется 36 молекул АТФ.
К некоторым гормонам и веществам на митохондриях имеются специализированные (аффинные) рецепторы. Трийодтиронин в норме ускоряет синтетическую активность митохондрий. Интерлейкин-1 и высокие концентрации трийодтиронина разобщают цепи окислительного фосфорилирования, вызывают набухание митохондрий, что сопровождается увеличением образования тепловой энергии.
Новые митохондрии образуются путем деления, перетяжкой или почкованием. В последнем случае образуется протомитохондрия, постепенно увеличивающаяся в размерах.
Протомитохондрия - мелкая органелла с наружной и внутренней мембранами. Внутренняя мембрана не имеет или содержит слаборазвитые кристы. Органелла характеризуется низким уровнем аэробного фосфорилирования. При образовании перетяжки содержимое митохондрии распределяется между двумя новыми довольно крупными органеллами. При любом способе размножения в каждой из вновь образующихся митохондрий имеется собственный геном.
Старые митохондрии разрушаются путем аутолиза (самопереваривания клеткой с помощью лизосом) с образованием аутолизосом. Из аутолизосомы образуется остаточное тельце. При полном переваривании содержимое остаточного тельца, состоящее из низкомолекулярных органических веществ, выводится путем экзоцитоза. При неполном переваривании остатки митохондрий могут накапливаться в клетке в виде слоистых телец или гранул с нипофусцином. В части митохондрий накапливаются нерастворимые соли кальция с образованием кристаллов - кальцинатов. Накопление продуктов дегенерации митохондрий может привести к дистрофии клетки.
Строение и функции митохондрий представляют собой довольно сложный вопрос. Наличие органеллы характерно почти для всех ядерных организмов – как для автотрофов (растений, способных к фотосинтезу), так и для гетеротрофов, которыми являются почти все животные, некоторые растения и грибы.
Главное предназначение митохондрий – окисление органических веществ и последующее использование освободившейся в результате этого процесса энергии. По этой причине органеллы имеют также и второе (неофициальное) название – энергетические станции клетки. Иногда их называют «пластидами катаболизма».
Что такое митохондрии
Термин имеет греческое происхождение. В переводе это слово означает «нить» (mitos), «зернышко» (chondrion). Митохондрии являются постоянными органоидами, которые имеют огромное значение для нормального функционирования клеток и делают возможным существование всего организма в целом.

«Станции» имеют специфическую внутреннюю структуру, которая изменяется в зависимости от функционального состояния митохондрии. Их форма может быть двух видов – овальная или продолговатая. Последняя нередко имеет ветвящийся вид. Число органоидов в одной клетке колеблется от 150 до 1500.
Особый случай – половые клетки. В сперматозоидах присутствует всего лишь одна спиральная органелла, в то время как женских гаметах содержится в сотни тысяч больше митохондрий. В клетке органоиды не зафиксированы в одном месте, а могут передвигаться по цитоплазме, совмещаться друг с другом. Их размер составляет 0,5 мкм, длина может достигать 60 мкм, в то время как минимальный показатель – 7 мкм.

Определить размер одной «энергетической станции» – непростая задача. Дело в том, что при рассмотрении в электронный микроскоп на срез попадает только часть органеллы. Случается так, что спиральная митохондрия имеет несколько сечений, которые можно принять за отдельные, самостоятельные структуры.
Только объемное изображение позволит выяснить точное клеточное строение и понять, идет речь о 2-5 отдельных органоидах или же об одной, имеющей сложную форму митохондрии.
Особенности строения
Оболочка митохондрии состоит из двух слоев: наружного и внутреннего. Последний включает в себя различные выросты и складки, которые имеют листовидную и трубчатую форму.
Каждая мембрана имеет особенный химический состав, определенное количество тех или иных ферментов и конкретное предназначение. Наружную оболочку от внутренней отделяет межмембранное пространство толщиной 10-20 нм.
Весьма наглядно выглядит строение органеллы на рисунке с подписями.

Схема строения митохондрии
Посмотрев на схему строения, можно сделать следующее описание. Вязкое пространство внутри митохондрии называется матриксом. Его состав создает благоприятную среду для протекания в ней необходимых химических процессов. В его составе присутствуют микроскопические гранулы, которые содействуют реакциям и биохимическим процессам (например, накапливают ионы гликогена и других веществ).
В матриксе находятся ДНК, коферменты, рибосомы , т-РНК, неорганические ионы. На поверхности внутреннего слоя оболочки располагаются АТФ-синтаза и цитохромы. Ферменты способствуют таким процессам, как цикл Кребса (ЦТК), окислительное фосфорилирование и т. д.
Таким образом, главная задача органоида выполняется как матриксом, так и внутренней стороной оболочки.
Функции митохондрий
Предназначение «энергетических станций» можно охарактеризовать двумя основными задачами:
- выработка энергии: в них осуществляются окислительные процессы с последующим выделением молекул АТФ;
- хранение генетической информации;
- участие в синтезе гормонов, аминокислот и других структур.
Процесс окисления и выработки энергии проходят в несколько стадий:


Схематичный рисунок синтеза АТФ
Стоит отметить: в результате цикла Кребса (цикл лимонной кислоты) не образуются молекулы АТФ, происходит окисление молекул и выделение углекислого газа. Это промежуточный этап между гликолизом и электронтранспортной цепью.
Таблица «Функции и строение митохондрий»

От чего зависит число митохондрий в клетке
Превалирующее число органоидов скапливается рядом с теми участками клетки, где возникает необходимость в энергетических ресурсах. В частности, большое количество органелл собирается в зоне нахождения миофибрилл, которые являются частью мышечных клеток, обеспечивающих их сокращение.

В мужских половых клетках структуры локализуются вокруг оси жгутика – предполагается, что потребность в АТФ обусловлена постоянным движением хвоста гаметы. Точно так же выглядит расположение митохондрий у простейших, которые для передвижения используют специальные реснички – органеллы скапливаются под мембраной у их основания.
Что касается нервных клеток, то локализация митохондрий наблюдается вблизи синапсов, через которые передаются сигналы нервной системы. В клетках, синтезирующих белки, органеллы скапливаются в зонах эргастоплазмы – они поставляют энергию, которая обеспечивает данный процесс.
Кто открыл митохондрии
Свое название клеточная структура обрела в 1897-1898 годах благодаря К. Бренду. Связь процессов клеточного дыхания с митохондриями сумел доказать Отто Вагбург в 1920 году.
Заключение
Митохондрии являются важнейшей составляющей живой клетки, выступая в роли энергетической станции, которая производит молекулы АТФ, обеспечивая тем самым процессы клеточной жизнедеятельности.
Работа митохондрий основана на окислении органических соединений, в результате чего происходит генерация энергетического потенциала.
Внешняя мембрана
Внутренняя мембрана
Матрикс
м-на, матрикс, кристы
. она имеет ровные контуры, не образует впячиваний или складок. На нее приходится около 7% от площади всех клеточных мембран. Ее толщина около 7 нм, она не бывает связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство
шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии,
ее матрикс или митоплазму. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист. Расстояние между мембранами в кристе составляет около 10-20 нм. Часто кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации. У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в нем иногда выявляются тонкие собранные в клубок нити (около 2-3 нм) и гранулы около 15-20нм. Теперь стало известно, что нити матрикса митохондрий представляют собой молекулы ДНК в составе митохондриального нуклеоида, а мелкие гранулы – митохондриальные рибосомы.
Функции митохондрий
1. В митохондриях происходит синтез ATP (см. Окислительное фосфорилирование)
PH межмембранного пространства ~4, pH матрикса ~8 | содержание белков в м: 67% - матрикс, 21% -наруж м-на, 6% - внутр м-на и 6% - в межм-ном пр-ве
Хандриома
– единая система митохондрий
наружная м-на: порины-поры позволяют проходить до 5 kD | внутренняя м-на: кардиолипин-делает непроницаемой м-ну для ионов |
межм-ное пр-во: группы ферментов фосфорилируют нуклеотиды и сахара нуклеотидов
внутренняя м-на:
матрикс: метаболические ферменты – окисление липидов, окисление углеводов, цикла трикарбоновых к-т, цикла Кребса
Происхождение от бактерий: амеба Pelomyxa palustris единств из эукариот не содержит м., живет в симбиозе с аэробными бактериями | собственная ДНК | схожие с бактериями оx процессы
Митохондриальная ДНК
Деление миохондрий
реплицируется
в интерфазе | репликация не связана с S-фазой | во время кл цикла митох один раз делятся надвое, образуя перетяжку, перетяжка сначала на внутр м-не | ~16,5 kb | кольцевая, кодирует 2 рРНК 22 тРНК и 13 белков |
транспорт белков: сигнальный пептид | амфифильный завиток | митохондриальный распознающий рецептор |
Окислительное фосфорилирование
Цепь переноса электронов
АТР-синтаза
в кл печени, м живут ~20 дней деление митохондрий путем образования перетяжки
16569пн=13белков,22тРНК,2pРНК | гладкая внешняя м-на (порины – проницаемость белков до 10 кДа) складчатая внутренняя (кристы) м-на (75% -белков: транспортные белки-переносчики, ф-ты, компаненты дыхат. цепи и АТФ-синтаза, кардиолипин) матрикс (обогащен ф-тами цитратного цикла) межм-ное пр-во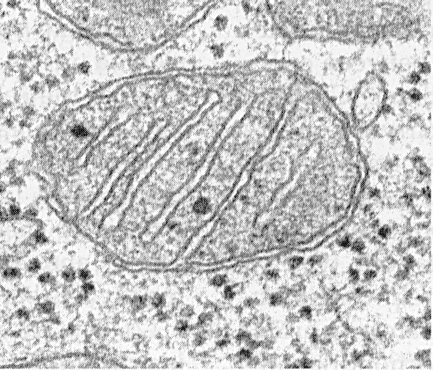
Митохондрии или хондриосомы (от греч. mitos - нить, chondrion - зернышко, soma - тельце) представляют собой гранулярные или нитевидные органеллы, присутствующие в цитоплазме простейших, растений и животных. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В живых клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом.
У разных видов размеры митохондрий очень непостоянны, так же как изменчива их форма (рис. 199). Все же у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм), а длина колеблется, достигая у нитчатых форм до 7-60 мкм.
Изучение величины и числа митохондрий не такое простое дело. Это связано с тем, что размеры и число митохондрий, которые видны на ультратонких срезах, не соответствуют реальности.
Обычные же подсчеты показывают, что на печеночную клетку приходится около 200 митохондрий. Это составляет более 20% от общего объема цитоплазмы и около 30-35% от общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны. Больше всего митохондрий в ооцитах (около 300000) и у гигантской амебы Chaos chaos (до 500000).
В клетках зеленых растений число митохондрий меньше, чем в клетках животных, так как часть их функций могут выполнять хлоропласты .
Локализация митохондрии в клетках различна. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы , где возникает потребность в АТФ, образующейся в митохондриях. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика; вероятно, это связано с необходимостью использования АТФ для движения хвоста сперматозоида. Аналогичным образом у простейших и в других клетках, снабженных ресничками, митохондрии локализуются непосредственно под клеточной мембраной у основания ресничек, для работы которых необходим АТФ. В аксонах нервных клеток митохондрии располагаются около синапсов, где происходит процесс передачи нервного импульса. В секреторных клетках, которые синтезируют большие количества белков, митохондрии тесно связаны с зонами эргастоплазмы; вероятно, они поставляют АТФ для активации аминокислот и синтеза белка на рибосомах .
Ультраструктура митохондрий.
Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами (рис. 205). Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы , она имеет ровные контуры, не образует впячиваний или складок, толщина составляет около 7 нм. На нее приходится около 7% от площади всех клеточных мембран. Мембрана не связана ни с какими другими мембранами цитоплазмы, замкнута сама на себя и представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс или митоплазму. Внутренняя мембрана митохондрий образовывает многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист.
Общая поверхность внутренней мембраны митохондрии в печеночной клетке составляет примерно треть поверхности всех клеточных мембран. Митохондрии клеток сердечной мышцы содержат втрое больше крист, чем печеночные митохондрии что отражает различия в функциональных нагрузках митохондрий разных клеток. Расстояние между мембранами в кристе составляет около 10-20 нм.
Митохондриальные кристы, отходящие от внутренней мембраны и простирающиеся в сторону матрикса, не перегораживают полностью полость митохондрии и не нарушают непрерывности заполняющего ее матрикса.
Ориентация крист по отношению к длинной оси митохондрии различна для разных клеток. Ориентация может быть перпендикулярная (клетки печени, почек) крист; в сердечной мышце наблюдается продольное расположение крист. Кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации (рис. 208). У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в котором выявляются молекулы ДНК в виде тонких собранных в клубок нитей (около 2-3 нм) и митохондриальные рибосомы имеющие форму гранул размером около 15-20 нм. Места отложения солей магния и кальция в матриксе образуют крупные (20-40 нм) плотные гранулы.
Функции митохондрий.
Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
Начальные этапы окисления углеводов называются анаэробным окислением, или гликолизом и происходят в гиалоплазме и не требуют участия кислорода. Субстратом окисления при анаэробном получении энергии служат гексозы и в первую очередь глюкоза; некоторые бактерии обладают свойством извлекать энергию, окисляяя пентозы, жирные кислоты или аминокислоты.
В глюкозе количество потенциальной энергии, заключенной в связях между атомами С, Н и О, составляет около 680 ккал на 1 моль (т.е. на 180 г глюкозы).
В живой клетке это огромное количество энергии освобождается в виде ступенчатого процесса, управляемого целым рядом окислительных ферментов, и не связано с переходом энергии химической связи в тепло, как при горении, а переходит в макроэнергетическую связь в молекулах АТФ, которые синтезируются при использовании освобождающейся энергии из АДФ и фосфата.
Образовавшиеся в результате гликолиза триозы, и в первую очередь пировиноградная кислота, в митохондриях вовлекаются в дальнейшее окисление. При этом происходит использование энергии расщепления всех химических связей, что приводит к выделению СО 2, к потреблению кислорода и синтезу большого количества АТФ. Эти процессы связаны с окислительным циклом трикарбоновых кислот и с дыхательной цепью переноса электронов, где происходит фосфорилирование АДФ и синтез клеточного “топлива”, молекул АТФ (рис. 209).
В цикле трикарбоновых кислот (цикл Кребса , или цикл лимонной кислоты) образовавшийся в результате гликолиза пируват сначала теряет молекулу СО 2 и, окисляясь до ацетата (двууглеродное соединение), соединяется с коферментом А. Затем ацетилкоэнзим А, соединяясь с оксалацетатом (четырехуглеродное соединение), образует шестиуглеродный цитрат (лимоную кислоту). Затем происходит цикл окисления этого шестиуглеродного соединения до четырехуглеродного оксалацетата, снова связывание с ацетилкоэнзимом А, и затем цикл повторяется. При этом окислении выделяются две молекулы СО 2, а электроны, освободившиеся при окислении, переносятся на акцепторные молекулы коферментов (NAD-никотинамидадениндинуклеотид), которые вовлекают их далее в цепь переноса электронов. Следовательно, в цикле трикарбоновых кислот нет самого синтеза АТФ, а идет окисление молекул, перенос электронов на акцепторы и выделение СО 2 . Все описанные выше события внутри митохондрий происходят в их матриксе.
Окисление исходного субстрата приводит к выделению СО 2 и воды, но при этом не выделяется тепловая энергия, как при горении, а образовываются молекулы АТФ. Они синтезируются другой группой белков, не связанных прямо с окислением. Во внутренних митохондриальных мембранах на поверхности мембран, смотрящих в матрикс, располагаются крупные белковые комплексы, ферменты, АТФ-синтетазы. В электронном микроскопе они видны в виде так называемых “грибовидных” телец сплошь выстилающие поверхность мембран, смотрящую в матрикс. Тельца имеют как бы ножку и головку, диаметром 8-9 нм. Следовательно, во внутренних мембранах митохондрий локализованы ферменты как окислительной цепи, так и ферменты синтеза АТФ (рис. 201б).
Дыхательная цепь - это главная система превращения энергии в митохондриях. Здесь происходит последовательное окисление и восстановление элементов дыхательной цепи, в результате чего высвобождается небольшими порциями энергия. За счет этой энергии в трех точках цепи из АДФ и фосфата образуется АТФ. Поэтому говорят, что окисление (перенос электронов) сопряжено с фосфорилированием (АДФ + Фн →АТФ, т.е. происходит процесс окислительного фосфорилирования.
Выделяющаяся при транспорте электронов энергия запасается в виде градиента протонов на мембране. Оказалось, что при переносе электронов в митохондриальной мембране каждый комплекс дыхательной цепи направляет свободную энергию окисление на перемещение протонов (положительных зарядов) через мембрану, из матрикса в межмембранное пространство, что приводит к образованию разности потенциалов на мембране: положительные заряды преобладают в межмембранном пространстве, а отрицательные - со стороны матрикса митохондрий. При достижении разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс, при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. Так происходит сопряжение окислительных процессов с синтетическим, с фосфорилированием АДФ. Пока происходит окисление субстратов, пока происходит перекачка протонов через внутреннюю митохондриальную мембрану - идет сопряженный с этим синтез АТФ, т.е. происходит окислительное фосфорилирование.
Эти два процесса можно разобщить. При этом продолжается перенос электронов, как и окисление субстрата, но синтеза АТФ не происходит. В этом случае энергия, освобождающаяся при окислении переходит в тепловую энергию.
Окислительное фосфорилирование у бактерий .
У прокариотических клеток, способных к окислительному фосфорилированию, элементы цикла трикарбоновых кислот локализованы прямо в цитоплазме, а ферменты дыхательной цепи и фосфорилирования связаны с клеточной мембраной, с ее выпячиваниями, выступающими внутрь цитоплазмы, с так называемыми мезосомами (рис. 212). Надо отметить, что такие бактериальные мезосомы могут быть связаны не только с процессами аэробного дыхания, но и у некоторых видов участвовать в делении клеток, в процессе распределения ДНК по новым клеткам, в образовании клеточной стенки и т.д.
На плазматической мембране в мезосомах некоторых бактерий осуществляются сопряженные процессы как окисления так и синтеза АТФ. В электронном микроскопе во фракциях плазматических мембран бактерий обнаружены сферические частицы, аналогичные тем, которые были найдены в митохондриях эукариотических клеток. Таким образом, у бактериальных клеток, способных к окислительному фосфорилированию, плазматическая мембрана выполняет роль, аналогичную внутренней мембране митохондрий эукариотических клеток.
Увеличение числа митохондрий.
Митохондрии могут увеличивать свою численность особенно при делении клеток или при увеличении функциональной нагрузки клетки. Происходит постоянное обновление митохондрий. Например, в печени средняя продолжительность жизни митохондрий составляет около 10 дней.
Увеличение числа митохондрий происходит путем роста и деления предшествующих митохондрий. Это предположение было впервые высказано Альтманом (1893), описавшим митохондрии под термином “биобласты”. Удается наблюдать прижизненно деление, фрагментацию длинных митохондрий на более короткие путем перетяжки, что напоминает бинарный способ деления бактерий.
Реальное увеличение числа митохондрий путем деления установлено при изучении поведения митохондрий в живых клетках культуры ткани. В течение клеточного цикла митохондрии вырастают до нескольких мкм, а затем фрагментируются, делятся на более мелкие тельца.
Митохондрии могут сливаться друг с другом и размножаться по принципу: митохондрия от митохондрии.
Авторепродукция митохондрий.
Двумембранные органеллы обладают полной системой авторепродукции. В митохондриях и пластидах существует ДНК, на которой синтезируются информационные, трансферные и рибосомные РНК и рибосомы, осуществляющие синтез митохондриальных и пластидных белков. Однако, эти системы, хотя и автономны, но ограничены по своим возможностям.
ДНК в митохондриях представляет собой циклические молекулы без гистонов и тем самым напоминают бактериальные хромосомы. Размер их составляет около 7 мкм, в одну циклическую молекулу митохондрий животных входит 16-19 тыс. нуклеотидных пар ДНК. У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представляют множественные копии, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре.
Так же как и у бактерий митохондральная ДНК собрана в отдельную зону - нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях может быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз в зависимости от типа клеток.
В некоторых культурах в клетках от 6 до 60% митохондрий не имеют нуклеоида, что может объясняться тем, что деление этих органелл скорее связано с фрагментацией, а не с распределением нуклеоидов.
Как уже говорилось, митохондрии могут как делиться, так и сливаться друг с другом. При слиянии митохондрий друг с другом может происходить обмен их внутренними компонентами.
Важно подчеркнуть, что рРНК и рибосомы митохондрий и цитоплазмы резко отличны. Если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s).
Рибосомные РНК митохондрий синтезируются на митохондриальных ДНК. В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий.
На митохондриальном геноме синтезируются 22 транспортные РНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х10 5 . В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х10 6 .
Если учесть, что кроме белков окислительного фосфорилирования в митохондрии входят ферменты цикла трикарбоновых кислот, ферменты синтеза ДНК и РНК, ферменты активации аминокислот и другие белки, то видно, что, для того чтобы кодировать эти многочисленные белки и рРНК и тРНК, количества генетической информации в короткой молекуле митохондриальной ДНК явно не хватает. Расшифровка нуклеотидной последовательности митохондриальной ДНК человека показала, что она кодирует всего лишь 2 рибосомные РНК, 22 трансферных РНК и всего 13 различных полипептидных цепей.
В настоящее время доказано, что большая часть белков митохондрий находится под генетическим контролем со стороны клеточного ядра и синтезируется вне митохондрий. Большинство митохондриальных белков синтезируется на рибосомах в цитозоле. Эти белки имеют специальные сигнальные последовательности, которые узнаются рецепторами на внешней мембране митохондрий. Эти белки могут встраиваться в них (см. аналогию с мембраной пероксисом), а затем перемещаться на внутреннюю мембрану. Этот перенос происходит в точках контакта наружной и внутренней мембран, где такой транспорт отмечен. Большинство липидов митохондрий так же синтезируются в цитоплазме.
Все это говорит о эндосимбиотическом происхождении митохондрий, о том, что митохондрии представляют собой организмы типа бактерий, находящиеся в симбиозе с эукариотический клеткой.
Хондриом.
Совокупность всех митохондрий в одной клетке называется хондриомом. Она может быть различной в зависимости от типа клеток. Во многих клетках хондриом состоит из разрозненных многочисленных митохондрий, равномерно расположенных по всей цитоплазме или локализуются группами в местах интенсивной траты АТФ. В обоих этих случаях митохондрии функционируют поодиночке, их кооперативная работа, возможно, координируется какими-то сигналами из цитоплазмы. Существует и совершенно иной тип хондриома, когда вместо мелких одиночных разрозненных митохондрий в клетке располагается одна гигантская разветвленная митохондрия.
Такие митохондрии встречаются у одноклеточных зеленых водорослей (например у Chlorella). Они образуют сложную митохондриальную сеть или митохондриальный ретикулум (Reticulum miyochondriale). Согласно хемоосмотической теории биологический смысл появления такой гигантской разветвленной митохондриальной структуры, объединенной в одно целое своими внешними и внутренними мембранами заключается в том, что в любой точке поверхности внутренней мембраны такой разветвленной митохондрии может идти синтез АТФ, который будет поступать в любую точку цитоплазмы, где в этом есть необходимость.
В случае гигантских разветвленных митохондрий в любой ее точке может на внутренней мембране накопиться потенциал, достаточный для того, чтобы начался синтез АТФ. С этих позиций митохондриальный ретикулум представляет собой как бы электрический проводник, кабель, соединяющий отдаленные точки такой системы. Митохондриальный ретикулум оказался очень полезным не только для мелких подвижных клеток, таких как хлорелла, но и для более крупных структурных единиц таких как, например, миофибриллы в скелетных мышцах.
Известно, что скелетные мышцы состоят из массы мышечных волокон, симпластов, содержащих множество ядер. Длина таких мышечных волокон достигает 40 мкм, при толщине 0,1 мкм - это гигантская структура, содержащая великое множество миофибрилл, все из которых сокращаются одновременно, синхронно. Для сокращения к каждой единице сокращения, к миофибрилле, доставляется большое количество АТФ, которых обеспечивают митохондрии на уровне z-дисков. На продольных ультратонких срезах скелетных мышц в электронном микроскопе видны многочисленные округлые мелкие сечения митохондрий, располагающихся в соседстве с саркомерами. Мышечные митохондрии представляют собой не мелкие шарики или палочки, а как бы паукообразные структуры, отростки которых ветвятся и простираются на большие расстояния, иногда через весь поперечник мышечного волокна.
При этом разветвления митохондрий окружают каждую миофибриллу в мышечном волокне, снабжая их АТФ, необходимого для мышечного сокращения. Следовательно, в плоскости z-диска митохондрии представляют типичный митохондриальный ретикулум. Такой пласт или этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных “поэтажных” пластов митохондриального ретикулума. Между “этажами” вдоль миофибрилл располагаются нитчатые митохондрии, соединяющие эти митохондриальные пласты. Таким образом создана трехмерная картина митохондриального ретикулума, проходящего через весь объем мышечного волокна.
Далее было установлено, что между ответвлениями митохондриального ретикулума и нитевидными продольными митохондриями существуют специальные межмитохондриальные соединения или контакты (ММК). Они образованы плотно прилегающими наружными митохондриальными мембранами контактирующих митохондрий, межмембранное пространство и мембраны в этой зоне имеют повышенную электронную плотность. Через эти специальные образования происходит функциональное объединение соседних митохондрий и митохондриальных ретикулумов в единую, кооперативную энергетическую систему. Все миофибриллы в мышечном волокне сокращаются синхронно по всей их длине, следовательно, и поступление АТФ на любом участке этой сложной машины тоже должно происходить синхронно, а это может происходить лишь в том случае, если огромное количество разветвленных митохондрий-проводников будет связано друг с другом с помощью контактов.
О том, что межмитоходриальные контакты (ММК) участвуют в энергетическом объединении митохондрий друг с другом удалось на кардиомиоцитах, клетках сердечных мышц.
Хондриом клеток сердечной мышцы не образует ветвящихся структур, а представлен множеством небольших вытянутых митохондрий, располагающихся без особого порядка между миофибриллами. Однако, все соседние митохондрии стыкуются друг с другом с помощью митохондриальных контактов такого же типа, как в скелетной мышце, только их число очень велико: в среднем на одну митохондрию приходится 2-3 ММК, которые связывают митохондрии в единую цепь, где каждым звеном такой цепи (Streptio mitochondriale) является отдельная митохондрия.
Оказалось, что межмитохондриальные контакты (ММК), как обязательная структура сердечных клеток обнаружены в кардиомиоцитах как желудочков, так и предсердий всех позвоночных животных: млекопитающих, птиц, пресмыкающихся, амфибий и костистых рыб. Более того ММК были обнаружены (но в меньшем числе) в клетках сердца некоторых насекомых и моллюсков.
Количество ММК в кардиомиоцитах изменяется в зависимости от функциональной нагрузки на сердце. Число ММК увеличивается при повышении физических нагрузок животных и, наоборот, при падении нагрузки на сердечную мышцу происходит резкое сокращение числа ММК.