Als innere Strukturen werden Mitochondrien bezeichnet. Die Struktur und Funktionen von Mitochondrien und Plastiden
Mitochondrien
In den Zellen tierischen Gewebes wurden Mitochondrien 1882 und in Pflanzen erst 1904 (in den Antheren von Seerosen) gefunden. Nach Auftrennung und Reinigung der Fraktion durch fraktionierte Zentrifugation konnten biologische Funktionen eingestellt werden. Sie enthalten 70% Protein und ca. 30% Lipide, eine geringe Menge RNA und DNA, Vitamine A, B 6, B 12, K, E, Fol- und Pantothensäure, Riboflavin, verschiedene Enzyme. Mitochondrien haben eine Doppelmembran, die äußere isoliert die Organelle vom Zytoplasma und die innere bildet Auswüchse von Crista. Der gesamte Raum zwischen den Membranen ist mit einer Matrix gefüllt (Abb. 13).
Die Hauptfunktion der Mitochondrien ist die Teilnahme an der Zellatmung. Die Rolle der Mitochondrien bei der Atmung wurde 1950-1951 festgelegt. Auf den Außenmembranen ist ein komplexes Enzymsystem des Krebszyklus konzentriert. Während der Oxidation von Atmungssubstraten wird Energie freigesetzt, die sich während der oxidativen Phosphorylierung in Kristallen sofort in den gebildeten ADP-Molekülen, hauptsächlich ATP, ansammelt. Die in den Makroverbindungen gespeicherte Energie wird künftig zur Deckung aller Bedürfnisse der Zelle verwendet.
Die Bildung von Mitochondrien in der Zelle erfolgt kontinuierlich aus den Mikrokörpern, häufiger ist ihr Auftreten mit der Differenzierung der Membranstrukturen der Zelle verbunden. Sie können in der Zelle durch Teilen und Knospen wiederhergestellt werden. Mitochondrien sind nicht dauerhaft, ihre Lebenserwartung beträgt 5-10 Tage.
Mitochondrien sind Kraftzellenstationen. Sie konzentrieren Energie, die in den "Akkumulatoren" der Energie - ATP - Moleküle gespeichert und nicht in der Zelle dissipiert wird. Eine Verletzung der Struktur der Mitochondrien führt zu einer Störung des Atmungsprozesses und infolgedessen zur Pathologie des Körpers.
Golgi-Apparat.Golgi-Apparat (synonym - dictiosomes) ist ein Stapel von 3-12 abgeflachten, geschlossenen, von einer Doppelmembran umgebenen Scheiben, Tanks genannt, an deren Rändern zahlreiche Blasen gelöst sind (300-500). Die Breite der Tanks beträgt 6-90 A. Die Dicke der Membranen beträgt 60-70 A.
Der Golgi-Apparat ist ein Zentrum für die Synthese, Akkumulation und Isolierung von Polysacchariden, insbesondere Cellulose, und ist an der Verteilung und dem intrazellulären Transport von Proteinen sowie an der Bildung von Vakuolen und Lysosomen beteiligt. In der Pflanzenzelle konnte die Beteiligung des Golgi-Apparats am Erscheinungsbild der Medianplatte und am Wachstum der Zellpekto-Cellulosemembran verfolgt werden.
Der Golgi-Apparat ist am weitesten in der Zeit des aktiven Zelllebens entwickelt. Mit zunehmendem Alter verkümmert es allmählich und verschwindet dann.
Lysosomen.Lysosomen - ziemlich kleine (ungefähr 0,5 Mikrometer im Durchmesser) gerundete Körper. Sie sind mit einer Protein-Lipoid-Membran beschichtet. Der Inhalt von Lysosomen sind zahlreiche hydrolytische Enzyme, die die Funktion des intrazellulären Aufschlusses (Lyse) von Proteinmakromolekülen, Nukleinsäuren, Polysacchariden erfüllen. Ihre Hauptfunktion ist die Verdauung einzelner Abschnitte des Zellprotoplasten (Autophagie - Selbstverschlingung). Dieser Prozess verläuft aufgrund von Phagozytose oder Pinozytose. Die biologische Rolle dieses Prozesses ist zweifach. Erstens ist es schützend, da bei einem vorübergehenden Mangel an Ersatzprodukten die Zelle aufgrund von Proteinen und anderen Substanzen das Leben unterstützt und zweitens überschüssige oder abgenutzte Organellen (Plastiden, Mitochondrien usw.) freigesetzt werden. Die Lysosomenmembran verhindert die Freisetzung von Enzymen in das Zytoplasma Andernfalls würde es von diesen Enzymen vollständig verdaut.
In einer toten Zelle werden Lysosomen zerstört, Enzyme sind in der Zelle und ihr gesamter Inhalt wird verdaut. Übrig bleibt nur die Pecto-Cellulose-Hülle.
Lysosomen sind Produkte der Aktivität des Golgi-Apparats, von ihm abgelöste Vesikel, in denen dieses Organoid Verdauungsenzyme ansammelt.
Spherosomen - abgerundete Protein-Lipoid-Körper von 0,3 bis 0,4 Mikron. Höchstwahrscheinlich handelt es sich um Derivate des Golgi-Apparats oder des endoplasmatischen Retikulums. In Form und Größe ähneln sie Lysosomen. Da Sphärosomen saure Phosphatase enthalten, sind sie wahrscheinlich mit Lysosomen verwandt. Einige Autoren glauben, dass Sphärosomen und Lysosomen einander äquivalent sind, höchstwahrscheinlich jedoch nur in Bezug auf Ursprung und Form. Es gibt eine Vermutung über ihre Beteiligung an der Synthese von Fetten (A. Frey-Wissling).
Ribosomen - sehr kleine Organoide mit einem Durchmesser von ca. 250 A. Sie sind fast kugelförmig. Einige von ihnen sind an die äußeren Membranen des endoplasmatischen Retikulums gebunden, einige von ihnen befinden sich im Zytoplasma in einem freien Zustand. Eine Zelle kann bis zu 5 Millionen Ribosomen enthalten. Ribosomen kommen in Chloroplasten und Mitochondrien vor, wo sie einen Teil der Proteine, aus denen diese Organoide bestehen, und die darin funktionierenden Enzyme synthetisieren.
Die Hauptfunktion ist die Synthese spezifischer Proteine \u200b\u200bnach Informationen aus dem Kern. Ihre Zusammensetzung: Protein und ribosomale Ribonukleinsäure (RNA) zu gleichen Anteilen. Ihre Struktur besteht aus kleinen und großen Untereinheiten, die aus Ribonukleotiden gebildet werden.
Mikrotubuli.Mikrotubuli - besondere Derivate des endoplasmatischen Retikulums. In vielen Zellen gefunden. Ihr Name selbst spricht von ihrer Form - ein oder zwei parallel angeordnete Röhrchen mit einem Hohlraum im Inneren. Außendurchmesser innerhalb von 250A. Die Wände der Mikrotubuli bestehen aus Proteinmolekülen. Spindelfäden werden während der Zellteilung aus Mikrotubuli gebildet.
Der Kern
Der Kern wurde 1831 von R. Brown in einer Pflanzenzelle entdeckt. Es befindet sich in der Mitte der Zelle oder in der Nähe der Zellmembran, ist aber allseitig von Zytoplasma umgeben. In den meisten Fällen befindet sich ein Kern in der Zelle, mehrere Kerne befinden sich in den Zellen einiger Algen sowie Pilze. Nichtzelluläre Grünalgen haben Hunderte von Kernen. Mehrkernige Zellen von nicht artikulierten Laktaturen. In den Zellen von Bakterien und Blaualgen befinden sich keine Kerne.
Die Form des Kerns kommt der Form einer Kugel oder Ellipse am häufigsten nahe. Hängt von der Form, dem Alter und der Funktion der Zelle ab. In der meristematischen Zelle ist der Kern groß, rund und nimmt 3/4 des Zellvolumens ein. In den Parenchymzellen der Epidermis mit einer großen zentralen Vakuole hat der Kern eine linsenförmige Form und wird zusammen mit dem Zytoplasma an die Peripherie der Zelle bewegt. Dies ist ein Zeichen für eine spezialisierte, aber bereits alternde Zelle. Eine kernlose Zelle kann nur kurze Zeit leben. Die kernfreien Zellen der Siebröhren sind lebende Zellen, leben aber nicht lange. In allen anderen Fällen sind kernfreie Zellen tot.
Der Kern hat eine Doppelschale, durch deren Poren der Inhalt gelangt
Kerne (Nukleoplasma) können mit dem Inhalt des Zytoplasmas kommunizieren. Die Membranen des Zellkerns sind mit Ribosomen ausgestattet und kommunizieren mit den Membranen des endoplasmatischen Retikulums der Zelle. Im Nukleoplasma befinden sich ein oder zwei Nukleolen und Chromosomen. Nucleoplasma ist ein kolloidales System von Zola, das in seiner Konsistenz einer verdickten Gelatine ähnelt. Nach Angaben russischer Biochemiker (Zbarsky I. B. et al.) Enthält der Kern vier Fraktionen von Proteinen: einfache Proteine \u200b\u200b- Globuline 20%, Desoxyribonukleoproteine \u200b\u200b- 70%, saure Proteine \u200b\u200b- 6% und Restproteine \u200b\u200b4%. Sie sind in folgenden Kernstrukturen lokalisiert: DNA-Proteide (alkalische Proteine) in den Chromosomen, RNA-Proteide (saure Proteine) in den Nukleolen, teilweise in den Chromosomen (bei der Synthese von Messenger-RNA) und in der Kernmembran. Globuline bilden die Basis des Nukleoplasmas. Restproteine \u200b\u200b(Natur nicht spezifiziert) bilden eine Kernmembran.
Der Großteil der Kernproteine \u200b\u200bsind komplexe alkalische Proteine, Desoxyribonukleoproteine, die auf DNA basieren.
DNA-Molekül.DNA-Molekül - Polynukleotid und besteht aus Nukleotiden. Das Nukleotid enthält drei Komponenten: ein Zuckermolekül (Desoxyribose), ein Stickstoffmolekül und ein Phosphorsäuremolekül. Desoxyribose ist mit der stickstoffhaltigen Base Glycosid und mit Phosphorsäure - einer Etherbindung - verbunden. In der DNA gibt es in verschiedenen Kombinationen nur 4 Arten von Nukleotiden, die sich in stickstoffhaltigen Basen voneinander unterscheiden. Zwei von ihnen (Adenin und Guanin) gehören zu stickstoffhaltigen Purinverbindungen und Cytosin und Thymin zu Pyrimidin. DNA-Moleküle befinden sich nicht in derselben Ebene, sondern bestehen aus zwei Spiralsträngen, d.h. Zwei parallele Ketten, die umeinander gewickelt sind, bilden ein DNA-Molekül. Sie sind durch Wasserstoffbrückenbindungen zwischen stickstoffhaltigen Basen miteinander verbunden, wobei die Purinbasen einer Kette mit den Pyrimidinbasen der anderen verbunden sind (Abb. 14). Die Struktur und Chemie des DNA-Moleküls wurde von englischen (Crick) und amerikanischen (Watson) Wissenschaftlern entdeckt und 1953 veröffentlicht. Dieser Punkt gilt als Beginn der Entwicklung der Molekulargenetik. Das Molekulargewicht der DNA beträgt 4–8 Mio. Die Anzahl der Nukleotide (verschiedene Optionen) beträgt bis zu 100.000. Das DNA-Molekül ist sehr stabil, seine Stabilität wird durch die Tatsache gewährleistet, dass es überall die gleiche Dicke aufweist - 20 Å (8 Å - die Breite der Pyrimidinbase + 12 Å - die Breite der Purinbase). Wenn radioaktiver Phosphor in den Körper eingeführt wird, wird die Markierung in allen phosphorhaltigen Verbindungen mit Ausnahme von DNA (Levy, Sickewitz) nachgewiesen.
DNA-Moleküle sind Vererbungsträger, weil Ihre Struktur kodiert Informationen über die Synthese spezifischer Proteine, die die Eigenschaften des Körpers bestimmen. Änderungen können unter dem Einfluss mutagener Faktoren (radioaktive Strahlung, starke chemische Agenzien, Alkaloide, Alkohole usw.) auftreten.
RNA-Molekül.Ribonukleinsäuremoleküle (RNA) deutlich weniger DNA-Moleküle. Dies sind einzelne Ketten von Nukleotiden. Es gibt drei Arten von RNA: Ribosomale, die längste, die zahlreiche Schleifen bildet, Information (Matrix) und Transport, die kürzeste. Ribosomale RNA ist in den Ribosomen des endoplasmatischen Retikulums lokalisiert und macht 85% der Gesamt-RNA der Zelle aus.
Informations-RNA ähnelt in ihrer Struktur einem Kleeblatt. Seine Menge beträgt 5% der Gesamt-RNA in der Zelle. Es wird in den Nukleolen synthetisiert. Seine Assemblierung erfolgt in Chromosomen während der Interphase. Seine Hauptfunktion ist die Übertragung von Informationen von der DNA auf die Ribosomen, wo die Proteinsynthese stattfindet.
Die Transport-RNA, wie sie jetzt etabliert ist, ist eine ganze Familie von Verbindungen, die in Struktur und biologischer Funktion verwandt sind. Nach einer groben Schätzung enthält jede lebende Zelle 40-50 einzelne Transport-RNAs, und ihre Gesamtzahl in der Natur ist angesichts der Artenunterschiede enorm. (Acad. V. Engelhardt). Sie werden Transport genannt, weil ihre Moleküle an der Aufrechterhaltung des Transports des intrazellulären Prozesses der Proteinsynthese beteiligt sind. In Kombination mit freien Aminosäuren liefern sie diese an die Ribosomen in der im Aufbau befindlichen Proteinkette. Dies sind die kleinsten RNA-Moleküle, die im Durchschnitt aus 80 Nukleotiden bestehen. Befindet sich in der zytoplasmatischen Matrix und macht etwa 10% der zellulären RNA aus
RNA enthält vier stickstoffhaltige Basen, aber im Gegensatz zu DNA ist Uracil in RNA anstelle von Thymin vorhanden.
Die Struktur von Chromosomen.Chromosomen wurden erstmals im späten 19. Jahrhundert von den Klassikern der Zytologie Fleming und Strasburger (1882, 1884) und dem russischen Zellforscher I.D. Tschistjakow entdeckte sie 1874.
Das Hauptstrukturelement der Chromosomen ist der Kern. Sie haben eine andere Form. Dies sind entweder gerade oder gebogene Stöcke, ovale Körper, Kugeln, deren Größe variiert.
Je nach Lage des Zentromers werden direkte, gleiche und ungleiche Chromosomen unterschieden. Die innere Struktur der Chromosomen ist in Abb. 1 dargestellt. Es ist anzumerken, dass das Desoxyribonukleoprotein ein Monomer des Chromosoms ist.
Im Chromosom der Desoxyribonukleoproteine \u200b\u200bsind 90-92%, davon 45% DNA und 55% Protein (Histon). RNA (informativ) ist auch in geringer Menge im Chromosom vertreten.
Das Chromosom hat auch eine ausgeprägte transversale Struktur - das Vorhandensein von verdickten Bereichen - Scheiben, die im Jahr 1909 waren. wurden Gene genannt. Dieser Begriff wurde vom dänischen Wissenschaftler Johansen vorgeschlagen. Der amerikanische Wissenschaftler Morgan bewies 1911, dass Gene die wichtigsten Erbeinheiten sind und in linearer Reihenfolge auf den Chromosomen verteilt sind. Daher weist das Chromosom qualitativ unterschiedliche Abschnitte auf. Der amerikanische Wissenschaftler Pinter hat 1934 die Diskontinuität der morphologischen Struktur von Chromosomen und das Vorhandensein von Scheiben in Chromosomen nachgewiesen. Scheiben sind Orte, an denen sich DNA ansammelt. Dies diente als Beginn der Erstellung von Chromosomenkarten, die den Ort (Locus) des Gens angaben, der das eine oder andere Merkmal des Organismus bestimmt. Ein Gen ist eine DNA-Doppelhelixregion, die Informationen über die Struktur eines einzelnen Proteins enthält. Dies ist ein Abschnitt eines DNA-Moleküls, der die Synthese eines einzelnen Proteinmoleküls bestimmt. DNA ist nicht direkt an der Proteinsynthese beteiligt. Es enthält und speichert nur Informationen über die Struktur des Proteins.
Die DNA-Struktur, bestehend aus mehreren tausend hintereinander angeordneten 4 Nukleotiden, ist ein Code der Vererbung.
Code der Vererbung. Proteinsynthese.Die erste Botschaft zum DNA-Code wurde 1961 vom amerikanischen Biochemiker Nirenberg in Moskau auf dem Internationalen Biochemischen Kongress verfasst. Das Wesen des DNA-Codes ist wie folgt. Jede Aminosäure entspricht einem Teil der DNA-Kette von drei benachbarten Nukleotiden (Tripletts). So entspricht beispielsweise eine Stelle, die aus TT-T (einem Triplett aus 3 Thymin-haltigen Nukleotiden) besteht, der Aminosäure Lysin, Triplett A (Adenin) -C (Cytosin) -A (Adenin) -Cystein usw. Angenommen, ein Gen wird durch eine Kette von Nukleotiden dargestellt, die in der folgenden Reihenfolge angeordnet sind: A-C-A-T-T-A-C-C-A-A-G-G. Nachdem wir diese Reihe in Tripletts zerlegt haben, werden wir sofort entschlüsseln, welche Aminosäuren und in welcher Reihenfolge sich im synthetisierten Protein befinden werden.
Die Anzahl der möglichen Kombinationen von 4 verfügbaren Nucleotiden von drei ist 4 × 64. Basierend auf diesen Verhältnissen wird die Anzahl der verschiedenen überschüssigen Tripletts ausreichen, um Informationen über die Synthese zahlreicher Proteine \u200b\u200bzu liefern, die sowohl die Struktur als auch die Funktionen des Körpers bestimmen. Für die Proteinsynthese wird eine exakte Kopie dieser Informationen in Form von Messenger-RNA an die Ribosomen gesendet. An der Entschlüsselung und Synthese sind neben i-RNA eine Vielzahl von Molekülen verschiedener Transport-Ribonukleinsäuren (t-RNAs), Ribosomen und einer Reihe von Enzymen beteiligt. Jede der 20 Aminosäuren bindet an T-RNA - ein Molekül mit einem Molekül. Jede der 20 Aminosäuren hat eine eigene t-RNA. T-RNAs haben chemische Gruppen, die in der Lage sind, ihre Aminosäure zu "erkennen", indem sie sie aus den verfügbaren Aminosäuren auswählen. Dies geschieht mit Hilfe spezieller Enzyme. Nachdem die t-RNA ihre Aminosäure erkannt hat, geht sie eine Verbindung mit ihr ein. Am Anfang der Kette (Molekül) der i-RNA befindet sich ein Ribosom, das auf seinem Weg entlang der i-RNA genau die Aminosäuren miteinander verbindet, deren Reihenfolge von der Nukleotidsequenz dieser I-RNA kodiert wird. Dadurch entsteht ein Proteinmolekül, dessen Zusammensetzung in einem der Gene kodiert ist.
Nucleoli- ein integraler struktureller Bestandteil des Kerns. Das sind kugelförmige Körper. Sie sind sehr wandelbar, verändern ihre Form und Struktur, erscheinen und verschwinden. Da sind eins, zwei. Für jede Pflanze eine bestimmte Anzahl. Die Kerne verschwinden, wenn sich die Zelle auf die Teilung vorbereitet, und erscheinen dann wieder. sie sind offensichtlich an der Synthese von Ribonukleinsäuren beteiligt. Wird der Nukleolus durch einen fokussierten Röntgen- oder Ultraviolettstrahl zerstört, wird die Zellteilung unterdrückt.
Die Rolle des Zellkerns im Zellleben. Der Zellkern dient als Kontrollzentrum der Zelle - er steuert die Zellaktivität und enthält Vererbungsträger (Gene), die die Eigenschaften eines bestimmten Organismus bestimmen. Die Rolle des Kerns kann identifiziert werden, wenn er mit mikrochirurgischen Techniken aus der Zelle entfernt und die Konsequenzen daraus beobachtet werden. Eine Reihe von Experimenten, die eine wichtige Rolle bei der Regulation des Zellwachstums nachweisen, wurde von Hemmerling an der einzelligen Grünalge Acetobularia durchgeführt. Dieser Seetang erreicht eine Höhe von 5 cm, sieht aus wie ein Pilz, hat die Ähnlichkeit von "Wurzeln" und "Beinen". Oben endet mit einem großen scheibenförmigen "Hut". Die Zelle dieser Alge hat einen Kern, der sich im basalen Teil der Zelle befindet.
Hammerling stellte fest, dass bei einem Beinschnitt das Unterteil weiterlebt und die Kappe nach der Operation vollständig regeneriert ist. Der kernlose obere Teil überlebt einige Zeit, stirbt aber am Ende ab, da er den unteren Teil nicht wiederherstellen kann. Folglich ist der Kern der Acetobularia für die dem Wachstum zugrunde liegenden Stoffwechselreaktionen notwendig.
Der Zellkern fördert die Zellwandbildung. Dies lässt sich durch Versuche mit der Alge Voucheria und Spyrogyra belegen. Durch die Freisetzung des Zellinhalts von geschnittenen Filamenten in Wasser können wir Zytoplasmaklumpen mit einem, mit mehreren Kernen und ohne Kerne erhalten. In den ersten beiden Fällen bildete sich die Zellmembran normal. In Abwesenheit eines Kerns bildete sich die Hülle nicht.
In den Experimenten von I. I. Gerasimov (1890) mit Spirogyra wurde gefunden, dass Zellen mit einem Doppelkern die doppelte Länge und Dicke des Chloroplasten aufweisen. In kernfreien Zellen setzt sich der Prozess der Photosynthese fort, es wird Assimilationsstärke gebildet, aber der Prozess ihrer Hydrolyse wird abgeschwächt, was durch das Fehlen von hydrolytischen Enzymen erklärt wird, die in Ribosomen nur gemäß der DNA des Kerns synthetisiert werden können. Das Leben eines Protoplasten ohne Kern ist minderwertig und kurzlebig. In den Experimenten von I.I. Gerasimovs kernfreie Spirogyra-Zellen lebten 42 Tage und starben. Eine der wichtigsten Funktionen des Zellkerns besteht darin, das Zytoplasma mit Ribonukleinsäure zu versorgen, die für die Proteinsynthese in der Zelle notwendig ist. Die Entfernung des Zellkerns aus der Zelle führt zu einer allmählichen Abnahme des RNA-Gehalts im Zytoplasma und einer Verlangsamung der Proteinsynthese darin.
Die wichtigste Rolle des Kerns ist die Übertragung von Merkmalen von Zelle zu Zelle, von Organismus zu Organismus, und zwar im Prozess der Kernspaltung und der Zelle als Ganzes.
Zellteilung.Zellen multiplizieren sich durch Division. In diesem Fall werden zwei Tochterzellen aus einer Zelle gebildet, wobei derselbe Satz von Erbmaterial in Chromosomen eingeschlossen ist wie die Mutterzelle. In somatischen Zellen werden Chromosomen durch zwei, die sogenannten homologen Chromosomen, dargestellt, in die Allelgene (Träger entgegengesetzter Zeichen, z. B. Blütenblätter von weißen und roten Erbsen usw.) eingelegt sind, Zeichen von zwei Elternpaaren. In dieser Hinsicht gibt es in den somatischen Zellen des Pflanzenkörpers immer einen doppelten Satz von Chromosen, der mit 2p bezeichnet wird. Chromosomen haben eine ausgeprägte Persönlichkeit. Die Quantität und Qualität der Chromosomen ist ein charakteristisches Merkmal jeder Art. In den Erdbeerzellen ist der diploide Chromosensatz 14 (2n), der Apfelbaum 34, die Topinambur 102 usw.
Mitose (Karyokinese) - Die somatische Zellteilung wurde erstmals von E. Russov (1872) und I. D. Chistyakov (1874) beschrieben. Die Essenz besteht darin, dass aus der Mutterzelle durch Teilung zwei Tochterzellen mit dem gleichen Chromosesatz gebildet werden, deren Zellzyklus sich aus der eigentlichen Interphase und Mitose zusammensetzt. Die Methode der Mikroautoradiographie stellte fest, dass die Interphase die längste und schwierigste ist - die Periode des "ruhenden" Kerns, weil In dieser Zeit kommt es zu einer Verdoppelung des Kernmaterials. Die Interphase ist in drei Phasen unterteilt:
Q1 - Presynthese (Dauer 4-6 Stunden);
S - synthetisch (10-20 Stunden);
Q2 - Postsynthese (2-5 Stunden).
Während der Phase Q1 laufen Vorbereitungen für die DNA-Reduktion. Und in der S-Phase wird DNA reduziert, die Zelle verdoppelt die DNA-Versorgung. In der Q2-Phase werden die zur Auslösung der Mitose notwendigen Enzyme und Strukturen gebildet. So werden in der Interphase DNA-Moleküle in Chromosomen in zwei identische Stränge aufgeteilt und Informations-RNA auf ihrer Matrix zusammengesetzt. Letzterer enthält Informationen über die Struktur spezifischer Proteine \u200b\u200bim Zytoplasma, und im Kern vervollständigt jeder der DNA-Stränge die fehlende Hälfte seines Moleküls. In diesem Prozess der Verdopplung (Reduktion) manifestiert sich ein einzigartiges Merkmal der DNA, das in der Fähigkeit der DNA besteht, sich selbst genau zu reproduzieren. Die resultierenden Tochter-DNA-Moleküle werden automatisch durch exakte Kopien des Elternmoleküls erhalten, weil Während der Reduktion verbinden sich die komplementären Basen (АТ; Г-Ц; usw.) aus der Umgebung zu jeder Hälfte.
In der Prophase der mitotischen Teilung machen sich doppelte Chromosomen bemerkbar. In der Metaphase befinden sie sich alle in der Äquatorzone in einer Reihe. Es bilden sich Spindelfäden (aus miteinander verbundenen Mikrotubuli). Die Hülle des Kerns und Nucleolus verschwinden. Verdickte Chromosomen werden der Länge nach in zwei Tochterchromosomen aufgeteilt. Dies ist das Wesen der Mitose. Es bietet eine genaue Verteilung der doppelten DNA-Moleküle zwischen den Tochterzellen. Dies stellt die Übertragung von Erbinformationen sicher, die in DNA verschlüsselt sind.
In der Anaphase beginnen sich Tochterchromosomen zu entgegengesetzten Polen zu bewegen. In der Mitte erscheinen die ersten Fragmente der Zellmembran (Fragmoblast).
In der Telophase werden in Tochterzellen Kerne gebildet. Der Inhalt der Mutterzelle (Organelle) verteilt sich auf die entstehenden Tochterzellen. Die Zellmembran ist vollständig ausgebildet. Damit ist die Zytokinese abgeschlossen (Abb. 17).
Abteilung Meiose - Reduktion wurde in den 90er Jahren des letzten Jahrhunderts von V. I. Belyaev entdeckt und beschrieben. Die Essenz der Teilung besteht darin, dass aus einer somatischen Zelle, die einen 2n-Chromosomensatz (doppelt, diploid) enthält, vier haploide Zellen gebildet werden, wobei "n" eine halbe Menge von Chromosen ist. Diese Art der Aufteilung ist komplex und besteht aus zwei Stufen. Die erste ist die Chromosomenreduktion. Doppelte Chromosomen befinden sich paarweise in der äquatorialen Zone (zwei parallele homologe Chromosomen). Zu diesem Zeitpunkt kann eine Konjugation (Adhäsion) durch Chromose, ein Crossover (Crossover) und infolgedessen ein Austausch von Chromosestellen auftreten. Infolgedessen wird ein Teil der Gene der väterlichen Chromosomen Teil der mütterlichen Chromosen und umgekehrt. Das Erscheinungsbild beider Chromosomen ändert sich dadurch nicht, ihre qualitative Zusammensetzung ändert sich jedoch. Die väterlichen und mütterlichen Vererbungen werden neu verteilt und gemischt.
In der Anaphase der Meiose divergieren homologe Chromosomen mit Hilfe von Spindelfäden an den Polen, an denen nach einer kurzen Ruhezeit (die Fäden verschwinden, aber das Septum zwischen den neuen Kernen bildet sich nicht) der Mitoseprozess einsetzt - eine Metaphase, in der alle Chromosomen in einer Ebene liegen und deren Längsspaltung stattfindet auf Tochterchromosomen. Bei einer anaphasischen Mitose mit Hilfe einer Spindel laufen sie an den Polen auseinander, wo sich vier Kerne und damit vier haploide Zellen bilden. In den Zellen einiger Gewebe tritt während ihrer Entwicklung unter dem Einfluss bestimmter Faktoren eine unvollständige Mitose auf, und die Anzahl der Chromosomen in den Kernen verdoppelt sich aufgrund der Tatsache, dass sie an den Polen nicht divergieren. Infolge solcher Verletzungen natürlicher oder künstlicher Natur entstehen Tetraploide und Polyploide. Mit Hilfe der Meiose werden Keimzellen gebildet - Gameten sowie Sporen, Elemente der sexuellen und asexuellen Vermehrung von Pflanzen (Abb. 18).
Amitose ist eine direkte Kernspaltung. Bei der Amitose bildet sich keine Teilungsspindel und die Hülle des Zellkerns zerfällt nicht wie bei der Mitose. Früher galt Amitose als primitive Form der Teilung. Es wurde nun festgestellt, dass es mit dem Abbau des Körpers verbunden ist. Es ist eine vereinfachte Version einer komplexeren Kernspaltung. Amitose tritt in den Zellen und Geweben von Nucellus, Endosperm, Knollenparenchym, Blattstielen usw. auf.
Mitochondrien - Es ist eine spiralförmige, runde, längliche oder verzweigte Organelle.
Das Konzept der Mitochondrien wurde erstmals 1897 von Bend vorgeschlagen. Mitochondrien können in lebenden Zellen mithilfe von Phasenkontrast- und Interferenzmikroskopie in Form von Körnern, Granulaten oder Filamenten nachgewiesen werden. Dies sind ziemlich mobile Strukturen, die sich bewegen, miteinander verschmelzen und teilen können. Mit speziellen Methoden in toten Zellen unter dem Lichtmikroskop gefärbt, wirken die Mitochondrien wie kleine Körner (Granulate), die im Zytoplasma diffus verteilt oder in bestimmten Zonen konzentriert sind.
Durch die Zerstörung von Glucose und Fetten in Gegenwart von Sauerstoff wird in den Mitochondrien Energie erzeugt und organische Substanz in Wasser und Kohlendioxid umgewandelt. Auf diese Weise erhalten tierische Organismen die für das Leben notwendige Grundenergie. Energie wird in Adenosintriphosphat (ATP) und genauer gesagt in seinen makroergischen Bindungen akkumuliert. Die Funktion der Mitochondrien hängt eng mit der Oxidation organischer Verbindungen und der Nutzung der bei ihrer Zersetzung freigesetzten Energie für die Synthese von ATP-Molekülen zusammen. Daher werden Mitochondrien oft als Zellenergiestationen oder Organellen der Zellatmung bezeichnet. ATP fungiert als Energielieferant, überträgt eine seiner energiereichen terminalen Phosphatgruppen auf ein anderes Molekül und wird zu ADP.
Es wird angenommen, dass Mitochondrien in der Evolution prokaryotische Mikroorganismen waren, die im Körper einer alten Zelle zu Symbiotika wurden. In der Folge wurden sie lebensnotwendig, was mit einem Anstieg des Sauerstoffgehalts in der Erdatmosphäre einherging. Einerseits haben Mitochondrien der Zelle überschüssigen giftigen Sauerstoff entzogen und andererseits Energie zugeführt.
Ohne Mitochondrien ist die Zelle praktisch nicht in der Lage, Sauerstoff als Energiequelle zu nutzen, und kann ihren Energiebedarf nur durch anaerobe Prozesse decken. Sauerstoff ist also Gift, aber Gift ist lebenswichtig für die Zelle, und überschüssiger Sauerstoff ist genauso schädlich wie sein Mangel.
Mitochondrien können ihre Form ändern und sich in die Bereiche der Zelle bewegen, in denen sie am dringendsten benötigt werden. In Kardiomyozyten befinden sich Mitochondrien in der Nähe von Myofibrillen, in den Zellen der Nierentubuli in der Nähe von basalen Vorsprüngen usw. Die Zelle enthält bis zu tausend Mitochondrien und ihre Anzahl hängt von der Aktivität der Zelle ab.
Mitochondrien haben eine durchschnittliche Quergröße von 0,5 ... 3 µm. Je nach Größe werden kleine, mittlere, große und riesige Mitochondrien unterschieden (sie bilden ein verzweigtes Netzwerk - das mitochondriale Retikulum). Die Größe und Anzahl der Mitochondrien hängen eng mit der Aktivität der Zelle und ihrem Energieverbrauch zusammen. Sie sind äußerst variabel und können je nach Zellaktivität, Sauerstoffgehalt und hormonellen Einflüssen anschwellen, die Anzahl und Struktur der Kristalle verändern, sich in Anzahl, Form und Größe sowie in der enzymatischen Aktivität unterscheiden.
Die Schüttdichte der Mitochondrien, der Entwicklungsgrad ihrer inneren Oberfläche und andere Indikatoren hängen vom Energiebedarf der Zelle ab. In den Lymphozyten gibt es nur wenige Mitochondrien und in den Leberzellen 2 ... 3 Tausend.
Mitochondrien bestehen aus einer Matrix, einer inneren Membran, einem perimitochondrialen Raum und einer äußeren Membran. Die äußere Mitochondrienmembran trennt die Organelle vom Hyaloplasma. Normalerweise hat es glatte Konturen und ist geschlossen, so dass es ein Membranbeutel ist.
Die äußere Membran ist von der inneren durch einen perimitochondrialen Raum mit einer Breite von ca. 10 ... 20 nm getrennt. Die innere Mitochondrienmembran begrenzt den tatsächlichen inneren Gehalt der Mitochondrien - der Matrix. Die innere Membran bildet zahlreiche Vorsprünge innerhalb der Mitochondrien, die das Aussehen von flachen Graten oder Kristallen haben.
Die Cristae können die Form von Platten (Trabekel) und Tubuli (Multivesicular am Schnitt) haben und sind in Bezug auf Mitochondrien in Längsrichtung oder in Querrichtung gerichtet.
Jede Mitochondrie ist mit einer Matrix gefüllt, die in elektronenmikroskopischen Aufnahmen dichter aussieht als das umgebende Zytoplasma. Die mitochondriale Matrix ist homogen (homogen), manchmal feinkörnig, mit verschiedenen Elektronendichten. Darin zeigen sich dünne Filamente mit einer Dicke von ca. 2 ... 3 nm und Granulate mit einer Größe von ca. 15 ... 20 nm. Matrixstränge sind DNA-Moleküle und kleine Körnchen sind mitochondriale Ribosomen. Die Matrix enthält Enzyme, eine einzelsträngige, cyclische DNA, mitochondriale Ribosomen und viele Ca 2+ -Ionen.
Das autonome System der Proteinsynthese von Mitochondrien wird durch histonfreie DNA-Moleküle dargestellt. DNA ist kurz, hat die Form eines Rings (zyklisch) und enthält 37 Gene. Im Gegensatz zur Kern-DNA gibt es darin praktisch keine nicht-kodierenden Nukleotidsequenzen. Merkmale der Struktur und Organisation bringen mitochondriale DNA mit bakterieller Zell-DNA zusammen. Auf der DNA der Mitochondrien findet eine Synthese von RNA-Molekülen verschiedener Arten statt: Information, Transfer (Transport) und ribosomale. Informations-RNA von Mitochondrien wird nicht gespleißt (Schnitte, die keine Informationslast tragen). Die geringe Größe der mitochondrialen DNA-Moleküle kann nicht die Synthese aller mitochondrialen Proteine \u200b\u200bbestimmen. Die meisten mitochondrialen Proteine \u200b\u200bstehen unter der genetischen Kontrolle des Zellkerns und werden im Zytoplasma synthetisiert, da mitochondriale DNA schwach exprimiert wird und nur einen Teil der oxidativen Phosphorylierungskettenenzyme bilden kann. Mitochondriale DNA kodiert nicht mehr als zehn Proteine, die in Membranen lokalisiert sind und Strukturproteine \u200b\u200bsind, die für die ordnungsgemäße Integration einzelner funktioneller Proteinkomplexe mitochondrialer Membranen verantwortlich sind. Proteine, die Transportfunktionen ausführen, werden ebenfalls synthetisiert. Ein solches System der Proteinsynthese bietet nicht alle Funktionen der Mitochondrien, daher ist die Autonomie der Mitochondrien begrenzt und relativ.
Bei Säugetieren werden Mitochondrien während der Befruchtung nur über die Eizelle übertragen, und Spermien bringen Nukleinsäure-DNA in den neuen Organismus ein.
In der Matrix der Mitochondrien werden Ribosomen gebildet, die sich von zytoplasmatischen Ribosomen unterscheiden. Sie sind an der Synthese einer Reihe von mitochondrialen Proteinen beteiligt, die nicht vom Zellkern kodiert werden. Mitochondriale Ribosomen haben eine Sedimentationszahl von 60 (im Gegensatz zu zytoplasmatischen Ribosomen mit einer Sedimentationszahl von 80). Die Sedimentationszahl ist die Abscheiderate während der Zentrifugation und Ultrazentrifugation. Mitochondriale Ribosomen sind strukturell den Ribosomen prokaryotischer Organismen ähnlich, jedoch kleiner und empfindlich gegenüber bestimmten Antibiotika (Chloramphenicol, Tetracyclin usw.).
Die innere Mitochondrienmembran weist eine hohe Selektivität beim Stofftransport auf. Die Enzyme der oxidativen Phosphorylierungskette, Elektronentransportproteine, ATP, ADP, Pyruvat und andere nahe beieinander liegende Transportsysteme sind an ihrer inneren Oberfläche gebunden Durch die enge Anordnung von Enzymen auf der inneren Membran wird eine hohe Konjugation (Interkonnektion) von biochemischen Prozessen gewährleistet, die die Geschwindigkeit erhöht und die Effizienz katalytischer Prozesse.
Elektronenmikroskopisch werden Pilzpartikel sichtbar, die in das Lumen der Matrix hineinragen. Sie haben ATP-synthetische Aktivität (bildet ATP aus ADP). Der Elektronentransport erfolgt entlang der in der inneren Membran lokalisierten Atmungskette, die vier große Enzymkomplexe (Cytochrome) enthält. Wenn Elektronen die Atmungskette passieren, werden Wasserstoffionen aus der Matrix in den perimitochondrialen Raum gepumpt, was die Bildung eines Protonengradienten (Pumpe) sicherstellt. Die Energie dieses Gradienten (Unterschiede in der Stoffkonzentration und der Bildung des Membranpotentials) wird zur ATP-Synthese und zum Transport von Metaboliten und anorganischen Ionen verwendet. Die in der inneren Membran enthaltenen Trägerproteine \u200b\u200btransportieren organische Phosphate, ATP, ADP, Aminosäuren, Fettsäuren, Tri- und Dicarbonsäuren durch diese.
Die äußere Mitochondrienmembran ist für niedermolekulare Substanzen durchlässiger, da sie viele hydrophile Proteinkanäle aufweist. Auf der Außenmembran befinden sich spezifische Rezeptorkomplexe, durch die Proteine \u200b\u200baus der Matrix in den perimitochondrialen Raum transportiert werden.
Die äußere Membran ist in ihrer chemischen Zusammensetzung und ihren Eigenschaften anderen intrazellulären Membranen und Plasmolemmen sehr ähnlich. Es enthält Enzyme, die Fette metabolisieren, die Umwandlung von Aminen und Aminooxidase aktivieren (katalysieren). Bleiben die Enzyme der Außenmembran aktiv, so ist dies ein Indikator für die Funktionssicherheit der Mitochondrien.
In den Mitochondrien gibt es zwei autonome Unterkompartimente. Da der perimitochondriale Raum oder die äußere Mitochondrienkammer (äußeres Teilkompartiment) aufgrund des Eindringens von Proteinkomplexen des Hyaloplasmas gebildet wird, wird das innere Teilkompartiment (mitochondriale Matrix) teilweise aufgrund der synthetischen Aktivität der mitochondrialen DNA gebildet. Das interne Teilkompartiment (Matrix) enthält DNA, RNA und Ribosomen. Es hat einen hohen Gehalt an Ca 2+ -Ionen im Vergleich zu Hyaloplasma. Im äußeren Teilraum sammeln sich Wasserstoffionen an. Die enzymatische Aktivität der äußeren und inneren Unterkompartimente, die Zusammensetzung der Proteine \u200b\u200bsind sehr unterschiedlich. Das innere Teilfach hat eine höhere Elektronendichte als das äußere.
Spezifische mitochondriale Marker sind die Enzyme Cytochromoxidase und Succinatdehydrogenase, deren Nachweis eine quantitative Charakterisierung von Energieprozessen in Mitochondrien ermöglicht.
Die Hauptfunktion der Mitochondrien - ATP-Synthese. Zunächst werden Zucker (Glucose) im Hyaloplasma zu Milchsäure und Brenztraubensäure (Pyruvat) unter gleichzeitiger Synthese einer geringen Menge ATP zerstört. Durch Glykolyse eines Glucosemoleküls werden zwei ATP-Moleküle verwendet und vier gebildet. Somit bilden nur zwei ATP-Moleküle die positive Bilanz. Diese Prozesse laufen ohne Sauerstoff ab (anaerobe Glykolyse).
Alle nachfolgenden Stufen der Energieerzeugung finden im Rahmen der aeroben Oxidation statt, bei der eine große Menge ATP synthetisiert wird. In diesem Fall wird organisches Material zu CO 2 und Wasser zerstört. Die Oxidation geht mit der Übertragung von Protonen auf ihre Akzeptoren einher. Diese Reaktionen werden mit einer Reihe von Enzymen des Tricarbonsäurezyklus durchgeführt, die sich in der Mitochondrienmatrix befinden.
Elektronentransfersysteme und die damit verbundene Phosphorylierung von ADP (oxidative Phosphorylierung) sind in die Kristallmembranen eingebaut. In diesem Fall erfolgt die Übertragung von Elektronen von einem Elektronenakzeptorprotein auf ein anderes und schließlich deren Bindung an Sauerstoff, wodurch Wasser gebildet wird. Gleichzeitig wird ein Teil der Energie, die bei einer solchen Oxidation in der Elektronentransferkette freigesetzt wird, während der ADP-Phosphorylierung als makroergische Bindung gespeichert, was zur Bildung einer großen Anzahl von ATP-Molekülen führt, dem wichtigsten intrazellulären Energieäquivalent. Der Prozess der oxidativen Phosphorylierung findet an den Membranen der Mitochondrienkristalle unter Verwendung der Oxidationskettenproteine \u200b\u200bund des hier befindlichen ADP-ATP-Synthetase-Phosphorylierungsenzyms statt. Durch oxidative Phosphorylierung entstehen aus einem Glucosemolekül 36 ATP-Moleküle.
Zu einigen Hormonen und Substanzen in den Mitochondrien gibt es spezialisierte (Affinitäts-) Rezeptoren. Triiodothyronin beschleunigt normalerweise die synthetische Aktivität von Mitochondrien. Interleukin-1 und hohe Konzentrationen von Triiodthyronin trennen die oxidativen Phosphorylierungsketten und verursachen eine mitochondriale Schwellung, die mit einer Zunahme der Bildung von Wärmeenergie einhergeht.
Neue Mitochondrien entstehen durch Teilung, Einschnürung oder Knospung. Im letzteren Fall werden Protomitochondrien gebildet, deren Größe allmählich zunimmt.
Protomitochondrien sind kleine Organellen mit äußeren und inneren Membranen. Die innere Membran hat oder enthält keine unterentwickelten Kristalle. Organella zeichnet sich durch eine geringe aerobe Phosphorylierung aus. Unter Bildung einer Einschnürung verteilt sich der Inhalt der Mitochondrien auf zwei neue, recht große Organellen. Bei jeder Fortpflanzungsmethode hat jede der neu gebildeten Mitochondrien ihr eigenes Genom.
Ältere Mitochondrien werden durch Autolyse (Selbstverdauung durch die Zelle unter Verwendung von Lysosomen) zerstört, um Autolysosomen zu bilden. Aus dem Autolysosom wird ein Restkörper gebildet. Bei vollständiger Verdauung wird der Restkörperinhalt, bestehend aus niedermolekularen organischen Substanzen, durch Exozytose ausgeschieden. Bei unvollständiger Verdauung können sich die Reste der Mitochondrien in Form von Schichtkörpern oder Granulaten mit Nipofuscin in der Zelle ansammeln. Unlösliche Calciumsalze reichern sich im Teil der Mitochondrien unter Bildung von Kristallen an - Verkalkungen. Die Anreicherung mitochondrialer Degenerationsprodukte kann zu Zelldystrophie führen.
Die Struktur und Funktion der Mitochondrien ist ein recht komplexes Thema. Das Vorhandensein von Organellen ist für fast alle Kernorganismen charakteristisch - sowohl für Autotrophen (Pflanzen, die zur Photosynthese fähig sind) als auch für Heterotrophen, bei denen es sich fast ausschließlich um Tiere, einige Pflanzen und Pilze handelt.
Hauptzweck der Mitochondrien ist die Oxidation organischer Substanzen und die anschließende Nutzung der dabei frei werdenden Energie. Aus diesem Grund haben Organellen auch einen zweiten (inoffiziellen) Namen - Zellenenergiestationen. Sie werden manchmal als "Katabolismus-Plastiden" bezeichnet.
Was ist Mitochondrien?
Der Begriff ist griechischen Ursprungs. Übersetzt bedeutet dieses Wort "Faden" (Mitos), "Samen" (Chondrion). Mitochondrien sind permanente Organoide, die für das normale Funktionieren von Zellen von großer Bedeutung sind und die Existenz des gesamten Organismus ermöglichen.

"Stationen" haben eine spezifische innere Struktur, die je nach Funktionszustand der Mitochondrien variiert. Ihre Form kann von zwei Arten sein - oval oder länglich. Letzteres hat oft ein verzweigtes Aussehen. Die Anzahl der Organellen in einer Zelle reicht von 150 bis 1500.
Ein Sonderfall sind Keimzellen. In den Spermien ist nur eine Spiralorganelle vorhanden, während die weiblichen Gameten Hunderttausende weiterer Mitochondrien enthalten. In der Zelle sind Organellen nicht an einem Ort fixiert, sondern können sich entlang des Zytoplasmas bewegen, sich miteinander verbinden. Ihre Größe beträgt 0,5 µm, die Länge kann 60 µm erreichen, während die Mindestgröße 7 µm beträgt.

Die Bestimmung der Größe eines „Kraftwerks“ ist keine leichte Aufgabe. Tatsache ist, dass bei der Untersuchung im Elektronenmikroskop nur ein Teil der Organelle in die Scheibe gelangt. Es kommt also vor, dass spiralförmige Mitochondrien mehrere Abschnitte aufweisen, die für separate, unabhängige Strukturen verwendet werden können.
Nur ein dreidimensionales Bild wird es uns ermöglichen, die genaue Zellstruktur herauszufinden und zu verstehen, ob es sich um 2-5 separate Organoide oder um eines mit einer komplexen Form von Mitochondrien handelt.
Strukturelle Merkmale
Die Mitochondrienmembran besteht aus zwei Schichten: außen und innen. Letzteres umfasst verschiedene Auswüchse und Falten, die eine blattartige und röhrenförmige Form haben.
Jede Membran hat eine spezielle chemische Zusammensetzung, eine bestimmte Menge bestimmter Enzyme und einen bestimmten Zweck. Die äußere Hülle ist von der inneren Hülle durch einen Zwischenmembranraum mit einer Dicke von 10 bis 20 nm getrennt.
Die Struktur der Organelle in der Abbildung mit den Bildunterschriften sieht sehr klar aus.

Die Struktur der Mitochondrien
Durch Betrachten des Strukturdiagramms kann die folgende Beschreibung vorgenommen werden. Der viskose Raum innerhalb der Mitochondrien wird als Matrix bezeichnet. Seine Zusammensetzung schafft ein günstiges Umfeld für den Fluss notwendiger chemischer Prozesse. Es enthält mikroskopisch kleine Granulate, die zu Reaktionen und biochemischen Prozessen beitragen (z. B. um Glykogenionen und andere Substanzen anzusammeln).
Die Matrix enthält DNA, Coenzyme, Ribosomen, t-RNA und anorganische Ionen. ATP-Synthase und Cytochrome befinden sich auf der Oberfläche der inneren Schicht der Schale. Enzyme tragen zu Prozessen wie dem Krebszyklus (CTK), der oxidativen Phosphorylierung usw. bei.
Somit wird die Hauptaufgabe des Organoids sowohl von der Matrix als auch von der Innenseite der Schale erfüllt.
Mitochondriale Funktion
Der Zweck der "Kraftwerke" kann durch zwei Hauptaufgaben charakterisiert werden:
- energieerzeugung: In ihnen werden Oxidationsprozesse mit anschließender Freisetzung von ATP-Molekülen durchgeführt;
- speicherung genetischer Informationen;
- beteiligung an der Synthese von Hormonen, Aminosäuren und anderen Strukturen.
Der Prozess der Oxidation und Energieerzeugung erfolgt in mehreren Schritten:


Schematische Darstellung der ATP-Synthese
Es ist erwähnenswert: Durch den Krebszyklus (Zitronensäurezyklus) werden keine ATP-Moleküle gebildet, die Moleküle oxidiert und Kohlendioxid freigesetzt. Dies ist ein Zwischenschritt zwischen der Glykolyse und der Elektronentransportkette.
Tabelle "Funktionen und Struktur der Mitochondrien"

Was bestimmt die Anzahl der Mitochondrien in einer Zelle
Die vorherrschende Anzahl von Organoiden sammelt sich in der Nähe der Teile der Zelle an, in denen ein Energiebedarf besteht. Insbesondere im Bereich der Myofibrillen wird eine große Anzahl von Organellen gesammelt, die Teil der Muskelzellen sind, die ihre Kontraktion sicherstellen.

In männlichen Keimzellen sind Strukturen um die Flagellenachse lokalisiert - es wird angenommen, dass der ATP-Bedarf auf die konstante Bewegung des Gametenschwanzes zurückzuführen ist. Die Anordnung der Mitochondrien in Protozoen, die spezielle Zilien für die Bewegung verwenden, sieht genauso aus, als würden sich Organellen unter der Membran an ihrer Basis ansammeln.
Bei Nervenzellen wird die Lokalisation von Mitochondrien in der Nähe der Synapsen beobachtet, durch die Signale des Nervensystems übertragen werden. In Zellen, die Proteine \u200b\u200bsynthetisieren, reichern sich Organellen in den Ergastoplasma-Zonen an - sie liefern Energie, die diesen Prozess liefert.
Wer entdeckte Mitochondrien
Die Zellstruktur erhielt ihren Namen in den Jahren 1897-1898 dank C. Brand. Otto Wagburg konnte 1920 den Zusammenhang der Zellatmung mit Mitochondrien nachweisen.
Fazit
Mitochondrien sind der wichtigste Bestandteil einer lebenden Zelle. Sie fungieren als Energiestation, die ATP-Moleküle produziert und so die Prozesse der zellulären Vitalaktivität sicherstellt.
Die Arbeit der Mitochondrien basiert auf der Oxidation organischer Verbindungen, wodurch Energiepotential erzeugt wird.
Äußere Membran
Innere Membran
Matrix m-on, matrix, krista. es hat glatte Konturen, bildet keine Vorsprünge oder Falten. Es macht etwa 7% der Fläche aller Zellmembranen aus. Seine Dicke beträgt ca. 7 nm, es ist nicht mit anderen Membranen des Zytoplasmas verbunden und für sich geschlossen, so dass es sich um einen Membranbeutel handelt. Die äußere Membran ist von der inneren getrennt zwischenmembranraum ungefähr 10-20 nm breit. Die innere Membran (ca. 7 nm dick) begrenzt den tatsächlichen Innengehalt der Mitochondrien.
seine Matrix oder Mitoplasma. Ein charakteristisches Merkmal der inneren Membran von Mitochondrien ist ihre Fähigkeit, zahlreiche Vorsprünge innerhalb der Mitochondrien auszubilden. Solche Vorsprünge haben meistens die Form von flachen Graten oder Kristallen. Der Abstand zwischen den Membranen in der Crista beträgt ca. 10-20 nm. Häufig können sich Kristalle in Form von Fingern verzweigen oder formen, biegen und weisen keine ausgeprägte Orientierung auf. In Protozoen, einzelligen Algen, in einigen Zellen höherer Pflanzen und Tiere sehen die Auswüchse der inneren Membran wie Röhren (Röhrenkristalle) aus.
Die mitochondriale Matrix hat eine feinkörnige homogene Struktur, manchmal werden dünne Filamente in einer Kugel (ca. 2-3 nm) gesammelt und Körnchen von ca. 15-20 nm darin nachgewiesen. Es ist nun bekannt geworden, dass mitochondriale Matrixstränge DNA-Moleküle in der Zusammensetzung des mitochondrialen Nukleoids sind und kleine Körnchen mitochondriale Ribosomen.
Mitochondriale Funktion
1. In Mitochondrien findet eine ATP-Synthese statt (siehe Oxidative Phosphorylierung)
Intermembran pH ~ 4, Matrix pH ~ 8 | Der Proteingehalt in m: 67% ist die Matrix, 21% ist die Entdeckung der Mine, 6% ist das Innere der Mine und 6% befindet sich in der Zwischenmine
Staupe - ein einziges mitochondriales System
äußere m-na: Porinporen ermöglichen den Durchtritt von bis zu 5 kD | inneres m-na: Cardiolipin-undurchlässiges m-well für Ionen |
Internationale Produktion: Enzymgruppen Phosphorylat-Nukleotide und Nukleotid-Zucker
Inland:
Matrix: metabolische Enzyme - Lipidoxidation, Kohlenhydratoxidation, Tricarbonsäurezyklus, Krebszyklus
Herkunft aus Bakterien: Die Amöbe Pelomyxa palustris enthält keine Einheit von Eukaryoten, lebt in Symbiose mit aeroben Bakterien eigene DNA | OX-ähnliche Bakterien
Mitochondriale DNA
Myochondriendivision
repliziert
in interphase | Die Replikation ist nicht mit der S-Phase verknüpft während des cl-Zyklus werden die Mitochis einmal zweigeteilt und bilden eine Einschnürung, wobei die Einschnürung zuerst auf der Innenseite erfolgt ~ 16,5 kb | Ring kodiert für 2 rRNA 22 tRNA und 13 Proteine \u200b\u200b|
Proteintransport: Signalpeptid | amphiphile Wellung mitochondrialer Erkennungsrezeptor
Oxidative Phosphorylierung
Elektronentransferschaltung
ATP-Synthase
In Leberzellen leben m ~ 20 Tage mitochondriale Teilung durch Bildung von Verengung
16569 bp \u003d 13 Proteine, 22tRNA, 2pRNA | glattes äußeres mn (porins - Durchlässigkeit von Proteinen bis 10 kDa) gefaltetes inneres (crista) mn (75% Proteine): Transportträgerproteine, f-you, Bestandteile der Atmungskette und ATP-Synthase, Cardiolipin) Matrix ( angereichert mit f-Tami-Citrat-Zyklus) inter 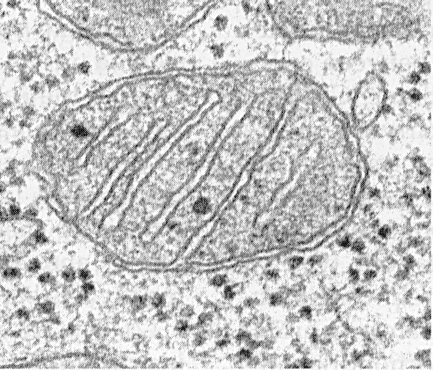
Mitochondrien oder Chondriosomen (aus dem Griechischen: Mitosfaden, Chondrionkorn, Somakörper) sind körnige oder filamentöse Organellen, die im Zytoplasma von Protozoen, Pflanzen und Tieren vorkommen. Mitochondrien können in lebenden Zellen beobachtet werden, da sie eine relativ hohe Dichte haben. In lebenden Zellen können sich Mitochondrien bewegen, bewegen und miteinander verschmelzen.
Bei verschiedenen Arten sind die Größen der Mitochondrien sehr unterschiedlich, ebenso wie ihre Form (Abb. 199). In den meisten Zellen ist die Dicke dieser Strukturen jedoch relativ konstant (etwa 0,5 μm), und die Länge schwankt und reicht in fadenförmigen Formen bis zu 7–60 μm.
Die Untersuchung der Größe und Anzahl der Mitochondrien ist nicht so einfach. Dies liegt daran, dass Größe und Anzahl der Mitochondrien, die auf ultradünnen Schnitten sichtbar sind, nicht der Realität entsprechen.
Herkömmliche Berechnungen zeigen, dass in der Leberzelle etwa 200 Mitochondrien vorhanden sind. Dies entspricht mehr als 20% des gesamten Zytoplasmas und etwa 30-35% der gesamten Proteinmenge in der Zelle. Die Oberfläche aller Mitochondrien der Leberzelle ist 4-5 mal größer als die Oberfläche ihrer Plasmamembran. Die meisten Mitochondrien befinden sich in Eizellen (ca. 300.000) und im Chaos der Riesenamöben (bis zu 500.000).
In grünen Pflanzenzellen ist die Anzahl der Mitochondrien geringer als in tierischen Zellen, da Chloroplasten einen Teil ihrer Funktionen erfüllen können.
Die Lokalisation der Mitochondrien in den Zellen ist unterschiedlich. Typischerweise reichern sich Mitochondrien in der Nähe der Bereiche des Zytoplasmas an, in denen ein Bedarf an ATP besteht, das in den Mitochondrien gebildet wird. Im Skelettmuskel befinden sich Mitochondrien in der Nähe von Myofibrillen. In Spermien bilden Mitochondrien eine Spirale um die Flagellenachse; Dies ist wahrscheinlich auf die Notwendigkeit zurückzuführen, ATP zu verwenden, um den Schwanz des Spermas zu bewegen. In Protozoen und anderen mit Zilien ausgestatteten Zellen befinden sich Mitochondrien direkt unter der Zellmembran an der Basis der Zilien, für deren Funktion ATP erforderlich ist. In den Axonen der Nervenzellen befinden sich Mitochondrien in der Nähe der Synapsen, wo der Prozess der Übertragung eines Nervenimpulses stattfindet. In sekretorischen Zellen, die große Mengen an Proteinen synthetisieren, sind Mitochondrien eng mit Ergastoplasma-Zonen verbunden. Sie liefern wahrscheinlich ATP, um Aminosäuren und die Proteinsynthese auf Ribosomen zu aktivieren.
Ultrastruktur der Mitochondrien.
Mitochondrien haben unabhängig von ihrer Größe oder Form eine universelle Struktur, ihre Ultrastruktur ist einheitlich. Mitochondrien sind auf zwei Membranen beschränkt (Abb. 205). Die äußere Mitochondrienmembran trennt sie vom Hyaloplasma, sie hat glatte Konturen, bildet keine Vorsprünge oder Falten und die Dicke beträgt etwa 7 nm. Es macht etwa 7% der Fläche aller Zellmembranen aus. Die Membran ist mit keiner anderen Membran des Zytoplasmas verbunden, ist in sich geschlossen und ist ein Membranbeutel. Die äußere Membran ist von der inneren durch einen Zwischenmembranraum mit einer Breite von etwa 10 bis 20 nm getrennt. Die innere Membran (ca. 7 nm dick) begrenzt den tatsächlichen inneren Gehalt der Mitochondrien, ihrer Matrix oder ihres Mitoplasmas. Die innere Mitochondrienmembran bildet zahlreiche Ausbuchtungen innerhalb der Mitochondrien. Solche Vorsprünge haben meistens die Form von flachen Graten oder Kristallen.
Die Gesamtoberfläche der inneren Mitochondrienmembran in der Leberzelle beträgt ungefähr ein Drittel der Oberfläche aller Zellmembranen. Die Mitochondrien der Herzmuskelzellen enthalten dreimal so viele Kristalle wie die Lebermitochondrien, was die unterschiedlichen funktionellen Belastungen der Mitochondrien verschiedener Zellen widerspiegelt. Der Abstand zwischen den Membranen in der Crista beträgt ca. 10-20 nm.
Mitochondriale Kristalle, die sich von der inneren Membran aus in Richtung der Matrix erstrecken, blockieren die mitochondriale Höhle nicht vollständig und beeinträchtigen nicht die Kontinuität der sie füllenden Matrix.
Die Orientierung der Kristalle in Bezug auf die Längsachse der Mitochondrien ist für verschiedene Zellen unterschiedlich. Die Ausrichtung kann senkrecht (Leber, Niere) sein. im Herzmuskel ist eine Längsanordnung von Kristallen zu beobachten. Crista kann sich fingerförmig verzweigen oder formen, sich biegen und keine ausgeprägte Orientierung haben (Abb. 208). In Protozoen, einzelligen Algen, in einigen Zellen höherer Pflanzen und Tiere sehen die Auswüchse der inneren Membran wie Röhren (Röhrenkristalle) aus.
Die mitochondriale Matrix weist eine feinkörnige homogene Struktur auf, in der DNA-Moleküle in Form von in einer Kugel gesammelten dünnen Filamenten (ca. 2-3 nm) und mitochondriale Ribosomen in Form von etwa 15-20 nm großen Granulaten nachgewiesen werden. Die Ablagerungen von Magnesium- und Calciumsalzen in der Matrix bilden große (20-40 nm) dichte Körnchen.
Die Funktionen der Mitochondrien.
Mitochondrien führen die ATP-Synthese durch, die durch Oxidation organischer Substrate und Phosphorylierung von ADP entsteht.
Die Anfangsstadien der Kohlenhydratoxidation werden als anaerobe Oxidation bezeichnet glykolyse und treten im Hyaloplasma auf und erfordern keine Beteiligung von Sauerstoff. Das Oxidationssubstrat während der anaeroben Energieerzeugung sind Hexosen und vor allem Glucose; Einige Bakterien haben die Fähigkeit, Energie durch Oxidation von Pentosen, Fettsäuren oder Aminosäuren zu extrahieren.
In Glucose beträgt die Menge an potentieller Energie, die in den Bindungen zwischen C-, H- und O-Atomen enthalten ist, etwa 680 kcal pro Mol (d. H. 180 g Glucose).
In einer lebenden Zelle wird diese enorme Energiemenge in Form eines schrittweisen Prozesses freigesetzt, der von einer Reihe oxidierender Enzyme gesteuert wird und nicht wie bei der Verbrennung mit dem Übergang der chemischen Bindungsenergie in Wärme verbunden ist, sondern in ATP-Molekülen, die unter Verwendung der freigesetzten Energie von in eine makroenergetische Bindung übergeht ADP und Phosphat.
Durch Glykolyse gebildete Triosen, hauptsächlich Brenztraubensäure, sind an der weiteren Oxidation in Mitochondrien beteiligt. In diesem Fall wird die Energie der Spaltung aller chemischen Bindungen genutzt, was zur Freisetzung von CO 2, zum Verbrauch von Sauerstoff und zur Synthese einer großen Menge ATP führt. Diese Prozesse sind mit dem Oxidationszyklus von Tricarbonsäuren und der Atmungskette des Elektronentransfers verbunden, bei dem die ADP-Phosphorylierung und die Synthese von zellulären „Brennstoff“ - und ATP-Molekülen stattfinden (Abb. 209).
Im Tricarbonsäure-Zyklus (Krebs-Zyklus oder Zitronensäure-Zyklus) verliert das durch Glykolyse gebildete Pyruvat zunächst das CO 2 -Molekül und verbindet sich bei Oxidation zu Acetat (einer 2-Kohlenstoff-Verbindung) mit Coenzym A. Anschließend verbindet sich Acetyl-Coenzym A mit Oxalacetat (einer 4-Kohlenstoff-Verbindung). bildet 6-Kohlenstoff-Citrat (Zitronensäure). Dann gibt es einen Oxidationszyklus dieser 6-Kohlenstoff-Verbindung zu 4-Kohlenstoff-Oxalacetat, der wiederum an Acetyl-Coenzym A bindet, und dann wiederholt sich der Zyklus. Während dieser Oxidation werden zwei CO 2 -Moleküle freigesetzt und die während der Oxidation freigesetzten Elektronen werden auf die Akzeptor-Coenzym-Moleküle (NAD-Nicotinamid-Adenin-Dinukleotid) übertragen, die sie ferner in die Elektronentransferkette einbeziehen. Daher gibt es im Kreislauf der Tricarbonsäuren keine ATP-Synthese selbst, sondern eine Oxidation von Molekülen, einen Elektronentransfer zu Akzeptoren und die Emission von CO 2. Alle oben beschriebenen Ereignisse in den Mitochondrien treten in ihrer Matrix auf.
Die Oxidation des Ausgangssubstrats führt zur Freisetzung von CO 2 und Wasser, jedoch wird wie bei der Verbrennung keine Wärmeenergie freigesetzt, sondern ATP-Moleküle gebildet. Sie werden von einer anderen Gruppe von Proteinen synthetisiert, die nicht in direktem Zusammenhang mit der Oxidation stehen. In den inneren Mitochondrienmembranen befinden sich große Proteinkomplexe, Enzyme und ATP-Synthetasen auf der Oberfläche der Membranen, die in die Matrix schauen. Im Elektronenmikroskop sind sie als sogenannte Pilzkörper sichtbar, die die Oberfläche der Membranen vollständig auskleiden und in die Matrix schauen. Die Körper haben ein Bein und einen Kopf mit einem Durchmesser von 8-9 nm. Folglich sind die Enzyme sowohl der Oxidationskette als auch der ATP-Synthese in den inneren Mitochondrienmembranen lokalisiert (Abb. 201b).
Die Atmungskette ist das wichtigste System zur Umwandlung von Energie in Mitochondrien. Hierbei kommt es zu einer sequentiellen Oxidation und Wiederherstellung der Elemente der Atmungskette, wodurch in geringen Anteilen Energie freigesetzt wird. Aufgrund dieser Energie wird ATP an drei Punkten in der Kette aus ADP und Phosphat gebildet. Daher wird gesagt, dass Oxidation (Elektronentransfer) mit Phosphorylierung (ADP + Fn → ATP) verbunden ist, d. H., Dass oxidative Phosphorylierung auftritt.
Die beim Elektronentransport freiwerdende Energie wird in Form eines Protonengradienten auf der Membran gespeichert. Es stellte sich heraus, dass während des Elektronentransfers in der Mitochondrienmembran jeder Komplex der Atmungskette die freie Oxidationsenergie leitet, um Protonen (positive Ladungen) durch die Membran von der Matrix in den Zwischenmembranraum zu bewegen, was zur Bildung von Potentialdifferenzen auf der Membran führt: im Zwischenmembranraum herrschen positive Ladungen, und negativ - von der Seite der mitochondrialen Matrix. Wenn die Potentialdifferenz (220 mV) erreicht ist, beginnt der Proteinkomplex der ATP-Synthetase, Protonen zurück in die Matrix zu transportieren und dabei eine Energieform in eine andere umzuwandeln: Er bildet ATP aus ADP und anorganischem Phosphat. Dies ist die Kombination von oxidativen Prozessen mit synthetischen unter Phosphorylierung von ADP. Während Substrate oxidieren, während Protonen durch die innere Mitochondrienmembran gepumpt werden, findet eine damit verbundene ATP-Synthese statt, d.h. es tritt eine oxidative Phosphorylierung auf.
Diese beiden Prozesse können getrennt werden. In diesem Fall wird der Elektronentransfer fortgesetzt, ebenso wie die Oxidation des Substrats, jedoch findet keine ATP-Synthese statt. In diesem Fall wird die bei der Oxidation freiwerdende Energie in Wärmeenergie umgewandelt.
Oxidative Phosphorylierung in Bakterien.
In prokaryontischen Zellen, die zur oxidativen Phosphorylierung befähigt sind, sind die Elemente des Tricarbonsäurezyklus direkt im Zytoplasma lokalisiert, und die Atmungsketten- und Phosphorylierungsenzyme sind mit der Zellmembran assoziiert, deren Vorsprünge in das Zytoplasma hineinragen, mit den sogenannten Mesosomen (Abb. 212). Es ist anzumerken, dass solche bakteriellen Mesosomen nicht nur mit den Prozessen der aeroben Atmung in Verbindung gebracht werden können, sondern auch bei einigen Arten an der Zellteilung, am Prozess der DNA-Verteilung in neuen Zellen, an der Bildung der Zellwand usw. beteiligt sind.
Auf der Plasmamembran in den Mesosomen einiger Bakterien werden konjugierte Prozesse sowohl der Oxidation als auch der ATP-Synthese durchgeführt. In den Fraktionen der Plasmamembranen von Bakterien wurden im Elektronenmikroskop kugelförmige Partikel gefunden, die denen in den Mitochondrien eukaryotischer Zellen ähnelten. Somit spielt in Bakterienzellen, die zur oxidativen Phosphorylierung fähig sind, die Plasmamembran eine ähnliche Rolle wie die innere Mitochondrienmembran von eukaryotischen Zellen.
Eine Zunahme der Anzahl der Mitochondrien.
Mitochondrien können ihre Anzahl insbesondere bei der Zellteilung oder bei einer Erhöhung der Funktionslast der Zelle erhöhen. Es gibt eine ständige Erneuerung der Mitochondrien. In der Leber beträgt die durchschnittliche Lebensdauer von Mitochondrien beispielsweise etwa 10 Tage.
Eine Zunahme der Anzahl der Mitochondrien erfolgt durch das Wachstum und die Teilung früherer Mitochondrien. Diese Annahme wurde erstmals von Altman (1893) geäußert, der Mitochondrien unter dem Begriff „Bioblasten“ beschrieb. Es ist möglich, in vivo eine Teilung, eine Fragmentierung langer Mitochondrien in kürzere durch Einschnürung, zu beobachten, die der binären Methode der Teilung von Bakterien ähnelt.
Eine reale Zunahme der Anzahl der Mitochondrien durch Teilung wurde festgestellt, als das Verhalten der Mitochondrien in lebenden Zellen der Gewebekultur untersucht wurde. Während des Zellzyklus wachsen Mitochondrien auf mehrere Mikrometer und teilen sich dann fragmentarisch in kleinere Körper auf.
Mitochondrien können miteinander verschmelzen und sich nach dem Prinzip vermehren: Mitochondrien aus Mitochondrien.
Autoproduktion von Mitochondrien.
Zwei-Membran-Organellen verfügen über ein vollständiges System der automatischen Reproduktion. In Mitochondrien und Plastiden gibt es DNA, auf der Informations-, Transfer- und Ribosomen-RNAs und Ribosomen synthetisiert werden, die die Synthese von Mitochondrien- und Plastidenproteinen durchführen. Diese Systeme sind zwar autonom, jedoch in ihren Fähigkeiten begrenzt.
DNA in Mitochondrien ist ein zyklisches Molekül ohne Histone und ähnelt daher bakteriellen Chromosomen. Ihre Größe beträgt etwa 7 Mikrometer, 16 bis 19.000 Nukleotid-DNA-Paare treten in ein zyklisches Molekül tierischer Mitochondrien ein. Beim Menschen enthält die mitochondriale DNA 16.500 Np. Und ist vollständig entschlüsselt. Es wurde festgestellt, dass die mitochondrale DNA verschiedener Objekte sehr homogen ist, ihr Unterschied liegt nur in der Größe der Introns und nicht transkribierten Stellen. Alle mitochondrialen DNAs sind mehrere Kopien, die in Gruppen und Clustern zusammengefasst sind. So können in einer Mitochondrie der Rattenleber 1 bis 50 cyclische DNA-Moleküle enthalten sein. Die Gesamtmenge an mitochondrialer DNA pro Zelle beträgt ungefähr ein Prozent. Die Synthese von mitochondrialer DNA hängt nicht mit der Synthese von DNA im Kern zusammen.
Genau wie bei Bakterien wird mitochondrale DNA in einer separaten Zone gesammelt - dem Nukleoid, dessen Größe etwa 0,4 Mikrometer im Durchmesser beträgt. In langen Mitochondrien können 1 bis 10 Nukleoide vorhanden sein. Bei der Teilung langer Mitochondrien wird ein Abschnitt, der ein Nukleoid enthält, von diesem getrennt (ähnlich der binären Teilung von Bakterien). Die DNA-Menge in einzelnen mitochondrialen Nukleoiden kann je nach Zelltyp 10-mal schwanken.
In einigen Kulturen haben in Zellen von 6 bis 60% der Mitochondrien kein Nukleoid, was durch die Tatsache erklärt werden kann, dass die Teilung dieser Organellen eher mit Fragmentierung als mit der Verteilung der Nukleoide verbunden ist.
Wie bereits erwähnt, können sich Mitochondrien teilen und miteinander verschmelzen. Wenn Mitochondrien miteinander verschmelzen, kann es zu einem Austausch ihrer inneren Bestandteile kommen.
Es ist wichtig zu betonen, dass sich rRNA und Ribosomen von Mitochondrien und Zytoplasma stark unterscheiden. Befinden sich im Zytoplasma 80s-Ribosomen, so gehören die mitochondrialen Ribosomen von Pflanzenzellen zu den 70s-Ribosomen (bestehen aus 30s und 50s-Untereinheiten, enthalten 16s und 23s-RNAs, die für prokaryotische Zellen charakteristisch sind), und kleinere Ribosomen werden in Mitochondrien tierischer Zellen gefunden (etwa 50s).
Mitochondriale ribosomale RNAs werden an mitochondrialer DNA synthetisiert. Im Mitoplasma auf den Ribosomen befindet sich die Proteinsynthese. Es stoppt im Gegensatz zur Synthese auf zytoplasmatischen Ribosomen unter der Wirkung des Antibiotikums Chloramphenicol, das die Proteinsynthese in Bakterien unterdrückt.
Auf dem Mitochondriengenom werden 22 Transport-RNAs synthetisiert. Der Triplettcode des mitochondrialen Synthesesystems unterscheidet sich von dem im Hyaloplasma verwendeten. Trotz des Vorhandenseins scheinbar aller für die Proteinsynthese erforderlichen Komponenten können kleine mitochondriale DNA-Moleküle nicht alle mitochondrialen Proteine, sondern nur einen kleinen Teil davon, kodieren. So DNA in der Größe von 15 Tausend n.p. können Proteine \u200b\u200bmit einem Gesamtmolekulargewicht von etwa 6 × 10 5 codieren. Gleichzeitig erreicht das Gesamtmolekulargewicht der Proteine \u200b\u200bder Partikel des gesamten Atmungsensembles der Mitochondrien einen Wert von etwa 2x10 6.
In Anbetracht der Tatsache, dass Mitochondrien neben oxidativen Phosphorylierungsproteinen auch Tricarbonsäurezyklusenzyme, DNA- und RNA-Syntheseenzyme, Aminosäureaktivierungsenzyme und andere Proteine \u200b\u200bumfassen, ist ersichtlich, dass die Menge an genetischer Information, um diese zahlreichen Proteine \u200b\u200bund rRNA und tRNA zu codieren, in das kurze mitochondriale DNA-Molekül fehlt eindeutig. Die Entschlüsselung der Nucleotidsequenz von menschlicher mitochondrialer DNA zeigte, dass sie nur 2 ribosomale RNAs, 22 Transfer-RNAs und insgesamt 13 verschiedene Polypeptidketten codiert.
Es ist jetzt nachgewiesen worden, dass die meisten mitochondrialen Proteine \u200b\u200bunter genetischer Kontrolle des Zellkerns stehen und außerhalb der Mitochondrien synthetisiert werden. Die meisten mitochondrialen Proteine \u200b\u200bwerden an Ribosomen im Cytosol synthetisiert. Diese Proteine \u200b\u200bhaben spezielle Signalsequenzen, die von Rezeptoren auf der äußeren Mitochondrienmembran erkannt werden. Diese Proteine \u200b\u200bkönnen in sie eingefügt werden (siehe Analogie zur Peroxisomenmembran) und dann zur inneren Membran wandern. Diese Übertragung erfolgt an den Kontaktstellen der äußeren und inneren Membranen, wo ein solcher Transport markiert ist. Die meisten mitochondrialen Lipide werden auch im Zytoplasma synthetisiert.
All dies weist auf den endosymbiotischen Ursprung von Mitochondrien hin, dh Mitochondrien sind Organismen wie Bakterien, die mit einer eukaryotischen Zelle in Symbiose stehen.
Chondriom.
Die Gesamtheit aller Mitochondrien in einer Zelle wird als Chondriom bezeichnet. Sie kann je nach Zelltyp unterschiedlich sein. In vielen Zellen bestehen Chondriome aus verstreuten Mitochondrien, die gleichmäßig im gesamten Zytoplasma verteilt oder in Gruppen an Orten mit intensivem ATP-Verbrauch lokalisiert sind. In beiden Fällen funktionieren Mitochondrien allein, ihre kooperative Arbeit wird möglicherweise durch einige Signale aus dem Zytoplasma koordiniert. Es gibt eine völlig andere Art von Chondriom, wenn sich anstelle von kleinen einzelnen verstreuten Mitochondrien eine riesige verzweigte Mitochondrie in der Zelle befindet.
Solche Mitochondrien kommen in einzelligen Grünalgen (z. B. Chlorella) vor. Sie bilden ein komplexes mitochondriales Netzwerk oder mitochondriales Retikulum (Reticulum miyochondriale). Nach der chemoosmotischen Theorie besteht die biologische Bedeutung des Auftretens einer solchen riesigen verzweigten Mitochondrienstruktur, die durch ihre äußeren und inneren Membranen verbunden ist, darin, dass die ATP-Synthese an jedem Punkt auf der Oberfläche der inneren Membran einer solchen verzweigten Mitochondrie stattfinden kann, der sich irgendwo im Zytoplasma befindet Dafür besteht Bedarf.
Bei riesig verzweigten Mitochondrien kann sich an jeder Stelle ein Potential an der inneren Membran ansammeln, das ausreicht, um die ATP-Synthese zu starten. Von diesen Positionen aus ist das mitochondriale Retikulum sozusagen ein elektrischer Leiter, ein Kabel, das die entfernten Punkte eines solchen Systems verbindet. Das mitochondriale Retikulum war nicht nur für kleine mobile Zellen wie Chlorella sehr nützlich, sondern auch für größere Struktureinheiten wie zum Beispiel Myofibrillen im Skelettmuskel.
Es ist bekannt, dass der Skelettmuskel aus einer Masse von Muskelfasern besteht, Symplasten, die viele Kerne enthalten. Die Länge solcher Muskelfasern erreicht 40 Mikrometer bei einer Dicke von 0,1 Mikrometern - dies ist eine gigantische Struktur, die sehr viele Myofibrillen enthält, die sich alle gleichzeitig synchron zusammenziehen. Zur Kontraktion wird eine große Menge ATP an jede Kontraktionseinheit, Myofibrille, abgegeben, die die Mitochondrien auf der Ebene der Z-Scheiben bereitstellen. Auf den ultradünnen Längsschnitten der Skelettmuskulatur zeigt ein Elektronenmikroskop zahlreiche abgerundete kleine Abschnitte von Mitochondrien in der Nähe von Sarkomeren. Muskuläre Mitochondrien sind keine kleinen Kugeln oder Stäbchen, sondern Spinnentierstrukturen, deren Prozesse sich über weite Strecken, manchmal über den gesamten Durchmesser der Muskelfaser, verzweigen und erstrecken.
Gleichzeitig umgibt eine mitochondriale Verzweigung jede Myofibrille in der Muskelfaser und versorgt sie mit dem für die Muskelkontraktion erforderlichen ATP. Daher stellen Mitochondrien in der Ebene der z-Scheibe ein typisches mitochondriales Retikulum dar. Eine solche Schicht oder ein solcher Boden des mitochondrialen Retikulums wird für jedes Sarkomer zweimal wiederholt, und die gesamte Muskelfaser weist Tausende von transversalen "Boden" -Schichten des mitochondrialen Retikulums auf. Zwischen den "Böden" entlang der Myofibrillen befinden sich filamentöse Mitochondrien, die diese Mitochondrienschichten verbinden. Auf diese Weise wurde ein dreidimensionales Bild des mitochondrialen Retikulums erstellt, das das gesamte Volumen der Muskelfaser durchquert.
Es wurde ferner festgestellt, dass zwischen den Zweigen des Mitochondrien-Retikulums und den filamentösen longitudinalen Mitochondrien spezielle intermitochondriale Verbindungen oder Kontakte (MMC) bestehen. Sie werden durch eng benachbarte äußere Mitochondrienmembranen gebildet, die mit Mitochondrien in Kontakt stehen, wobei der Zwischenmembranraum und die Membranen in dieser Zone eine erhöhte Elektronendichte aufweisen. Durch diese speziellen Formationen findet eine funktionelle Vereinigung benachbarter Mitochondrien und mitochondrialer Retikula zu einem einzigen kooperativen Energiesystem statt. Alle Myofibrillen in der Muskelfaser werden synchron über ihre gesamte Länge reduziert, daher sollte die ATP-Zufuhr in jedem Teil dieser komplexen Maschine auch synchron erfolgen, und dies kann nur geschehen, wenn eine große Anzahl von verzweigten Mitochondrienleitern miteinander verbunden sind Kontakte verwenden.
Die Tatsache, dass intermitochterielle Kontakte (MMK) an der Energie-Assoziation von Mitochondrien beteiligt sind, war auf Kardiomyozyten, Herzmuskelzellen, möglich.
Chondriome von Herzmuskelzellen bilden keine Verzweigungsstrukturen, sondern werden durch viele kleine längliche Mitochondrien dargestellt, die sich ohne besondere Ordnung zwischen den Myofibrillen befinden. Jedoch verbinden sich alle benachbarten Mitochondrien mit Hilfe von Mitochondrienkontakten des gleichen Typs wie im Skelettmuskel, nur ist ihre Anzahl sehr groß: Im Durchschnitt gibt es 2-3 MMCs pro Mitochondrien, die Mitochondrien in einer einzelnen Kette binden, wobei jedes Glied hat Kette (Streptio mitochondriale) ist eine separate Mitochondrien.
Es stellte sich heraus, dass intermitochondriale Kontakte (MMC) als obligatorische Struktur von Herzzellen in Kardiomyozyten sowohl der Ventrikel als auch der Vorhöfe aller Wirbeltiere gefunden wurden: Säugetiere, Vögel, Reptilien, Amphibien und Knochenfische. Darüber hinaus wurden MMCs (jedoch in geringerer Anzahl) in den Herzzellen einiger Insekten und Weichtiere gefunden.
Die Menge an MMC in Kardiomyozyten variiert in Abhängigkeit von der funktionellen Belastung des Herzens. Die Zahl der MMC nimmt mit zunehmender körperlicher Aktivität der Tiere zu, und umgekehrt nimmt die Zahl der MMC stark ab, wenn die Belastung des Herzmuskels abnimmt.







