Kako se imenujejo notranje strukture mitohondrijev? Zgradba in funkcija mitohondrijev in plastid
Mitohondrije
V celicah živalskih tkiv so mitohondrije našli leta 1882, v rastlinah pa - šele leta 1904 (v prašicah vodne lilije). Biološke funkcije so bile določene po izolaciji in čiščenju frakcije s frakcijskim centrifugiranjem. Vsebujejo 70% beljakovin in približno 30% lipidov, majhno količino RNK in DNK, vitamine A, B 6, B 12, K, E, folno in pantotensko kislino, riboflavin, različne encime. Mitohondriji imajo dvojno membrano, zunanja izolira organelo iz citoplazme, notranja pa tvori izrastke kriste. Celoten prostor med membranami je napolnjen z matrico (slika 13).
Glavna funkcija mitohondrijev je sodelovanje v celičnem dihanju. Vloga mitohondrijev pri dihanju je bila ugotovljena v letih 1950-1951. Kompleksni encimski sistem Krebsovega cikla je koncentriran na zunanjih membranah. Ko se substrati dihanja oksidirajo, se sprosti energija, ki se takoj v procesu oksidativne fosforilacije, ki se pojavi v kristalih, nabira v tvorjenih molekulah ADP in večinoma ATP. Energija, shranjena v visokoenergetskih spojinah, se v prihodnosti uporablja za zadovoljevanje vseh potreb celice.
Nastajanje mitohondrijev v celici poteka neprestano iz mikroteles, pogosteje je njihov nastanek povezan z diferenciacijo struktur celične membrane. V celici jih je mogoče obnoviti tako, da jih delimo in poparimo. Mitohondriji niso trpežni, njihova življenjska doba je 5-10 dni.
Mitohondrije so "napajalne" postaje celice. Koncentrirajo energijo, ki je shranjena v energijskih "akumulatorjih" - molekulah ATP, in se ne raztrese v celici. Kršitev mitohondrijske strukture vodi do motenj dihalnega procesa in posledično do patologije telesa.
Golgijev aparat.Golgijev aparat (sinonim - dictyosomi) je kup 3-12 sploščenih, zaprtih, obdan z dvojno membrano diskov, imenovane cisterne, od robov katerih so odcepljeni številni vezikli (300–500). Širina rezervoarjev je 6-90 A, debelina membran 60-70 A.
Golgijev aparat je središče za sintezo, kopičenje in sproščanje polisaharidov, zlasti celuloze, sodeluje pri porazdelitvi in \u200b\u200bmedceličnem transportu beljakovin, pa tudi pri tvorbi vakuolov in lizosomov. V rastlinski celici je bilo mogoče izslediti udeležbo Golgijevega aparata pri nastajanju mediane plošče in rasti celične pekto-celulozne membrane.
Golgijev aparat je najbolj razvit v obdobju aktivnega celičnega življenja. S staranjem se postopoma atrofira, nato pa izgine.
Lizosomi.Lizosomi - precej majhna (približno 0,5 mikrona v premeru) zaobljena telesa. Prekrite so z beljakovinsko-lipoidno membrano. Vsebine lizosomov so številni hidrolizni encimi, ki opravljajo funkcijo znotrajcelične prebave (lize) beljakovinskih makromolekul, nukleinskih kislin, polisaharidov. Njihova glavna funkcija je prebava posameznih odsekov celičnega protoplasta (avtofagija - samopotrošnja). Ta proces poteka skozi fagocitozo ali pinocitozo. Biološka vloga tega procesa je dvojna. Prvič, zaščitna je, saj z začasnim pomanjkanjem izdelkov za shranjevanje celica vzdržuje življenje zaradi ustavnih beljakovin in drugih snovi, in drugič, pride do odvajanja odvečnih ali dotrajanih organelov (plastid, mitohondrije itd.) Lizosomna ovojnica preprečuje sproščanje encimov v citoplazmi. sicer bi jih vsi encimi prebavili.
V umrli celici se lizosomi uničijo, v celici najdemo encime in prebavimo vso njeno vsebino. Ostane le še pekto-celulozna lupina.
Lizosomi so produkti Golgijevega aparata, iz njega odtrgani mehurčki, v katerih je ta organoid nabral prebavljive encime.
Sferosomi - zaobljena beljakovinsko-lipoidna telesa 0,3-0,4 mikrona. Po vsej verjetnosti so derivati \u200b\u200bGolgijevega aparata ali endoplazemskega retikuluma. Po obliki in velikosti spominjajo na lizosome. Ker sferosomi vsebujejo kislo fosfatazo, so verjetno povezani z lizosomi. Nekateri avtorji menijo, da so sferosomi in lizosomi med seboj enakovredni, vendar najverjetneje le po izvoru in obliki. Obstaja domneva o njihovem sodelovanju pri sintezi maščob (A. Frey-Wissling).
Ribosomi - zelo majhne organele, njihov premer je približno 250A, so skoraj kroglaste oblike. Nekateri od njih so pritrjeni na zunanje membrane endoplazemskega retikuluma, nekateri so v citoplazmi v prostem stanju. Celica lahko vsebuje do 5 milijonov ribosomov. Ribosome najdemo v kloroplastih in mitohondrijih, kjer sintetizirajo nekatere beljakovine, iz katerih so zgrajene te organele, in encime, ki delujejo v njih.
Glavna funkcija je sinteza specifičnih beljakovin glede na informacije, ki prihajajo iz jedra. Njihova sestava: beljakovine in ribosomalna ribonukleinska kislina (RNA) v enakih razmerjih. Njihova struktura je majhna in velika podenota, ki nastane iz ribonukleotida.
Mikrotubule.Mikrotubule - neke vrste derivatov endoplazemskega retikuluma. Najdemo ga v številnih celicah. Že njihovo ime govori o njihovi obliki - ena ali dve vzporedni cevi s votlino v notranjosti. Zunanji premer znotraj 250A. Stene mikrotubul so zgrajene iz molekul beljakovin. Vretenasti niti se tvorijo iz mikrotubul med celično delitvijo.
Jedro
Jedro je leta 1831 v rastlinski celici odkril R. Brown. Nahaja se v središču celice ali blizu celične membrane, vendar ga na vseh straneh obdaja citoplazma. V večini primerov je v celici eno jedro, več jeder je v celicah nekaterih alg, pa tudi gliv. Zelene alge necelične strukture imajo na stotine jeder. Večjedrne celice nesegmentiranih laktatov. V celicah bakterij in modro-zelenih alg ni jeder.
Oblika jedra je najpogosteje blizu oblike kroglice ali elipse. Odvisno je od oblike, starosti in funkcije celice. V meristematski celici je jedro veliko, zaobljeno in zavzema 3/4 volumna celice. V parenhimskih celicah povrhnjice, ki imajo veliko osrednjo vakuolo, ima jedro lečasto obliko in se potisne skupaj s citoplazmo na obod celice. To je znak specializirane, a že senescentne celice. Celica, ki je brez jedra, lahko živi le kratek čas. Celice brez jedra sitov so žive celice, vendar ne živijo dolgo. V vseh drugih primerih so nejedrske celice mrtve.
Jedro ima dvojno lupino, skozi pore, v katerih je vsebina
jedro (nukleoplazma) lahko komunicira z vsebino citoplazme. Mreže membrane jedra so opremljene z ribosomi in komunicirajo z membranami endoplazemskega retikuluma celice. Eden ali dva nukleola in kromosomi se nahajajo v nukleoplazmi. Nukleoplazma je koloidni sistem sol, ki po konsistenci spominja na odebeljeno želatino. Jedro po mnenju domačih biokemikov (Zbarskiy I.B. et al.) Vsebuje štiri frakcije beljakovin: enostavne beljakovine - globulini 20%, deoksiribonukleoproteini - 70%, kisle beljakovine - 6% in preostale beljakovine 4%. Lokalizirani so v naslednjih jedrskih strukturah: proteini DNA (alkalni proteini) - v kromosomih, proteini RNA (kisli proteini) - v nukleolih, delno v kromosomih (med sintezo selitvene RNA) in v jedrski membrani. Globulini tvorijo osnovo nukleoplazme. Preostali proteini (narava ni določena) tvorijo jedrsko membrano.
Večji del jedrskih beljakovin predstavljajo kompleksni alkalni proteini, deoksiribonukleoproteini, ki temeljijo na DNK.
Molekula DNA.Molekula DNA - polinukleotid in je sestavljen iz nukleotidov. Nukleotid vsebuje tri komponente: molekulo sladkorja (deoksiriboza), molekulo dušikove baze in molekulo fosforjeve kisline. Deoksiriboza je povezana z dušikovo bazo glikozida, fosforna kislina pa z etrsko vezjo. V DNK so samo 4 vrste nukleotidov v različnih kombinacijah, ki se med seboj razlikujejo po dušikovih bazah. Dva izmed njih (adenin in gvanin) sta dušikovi purinski spojini, citozin in timin pa pirimidinske spojine. Molekule DNK so nameščene ne v eni ravnini, ampak so sestavljene iz dveh navitih pramenov, tj. dve vzporedni verigi, zviti okoli druge, tvorita eno molekulo DNK. Med seboj se vežejo z vodikovo vezjo med dušikovimi bazami, purinske baze ene verige pa pritrjujejo pirimidinske baze druge (slika 14). Strukturo in kemijo molekule DNA so angleški znanstveniki (Crick) in ameriški (Watson) odkrili in objavili leta 1953. Ta trenutek velja za začetek razvoja molekularne genetike. Molekularna teža DNK je 4-8 milijonov. Število nukleotidov (različne variante) je do 100 tisoč. Molekula DNA je zelo stabilna, njeno stabilnost zagotavlja dejstvo, da ima v celotni dolžini enako debelino - 20A (8A - širina baze pirimidina + 12A - širina baze purina). Če v telo vnesemo radioaktivni fosfor, potem najdemo etiketo v vseh spojinah, ki vsebujejo fosfor, razen v DNK (Levy, Sikewitz).
Molekule DNK so nosilci dednosti, ker njihova struktura kodira podatke o sintezi specifičnih beljakovin, ki določajo lastnosti organizma. Spremembe se lahko pojavijo pod vplivom mutagenih dejavnikov (radioaktivno sevanje, močna, kemična sredstva - alkaloidi, alkoholi itd.).
Molekula RNA.Molekule ribonukleinske kisline (RNA) bistveno manj molekul DNK. To so enojne verige nukleotidov. Obstajajo tri vrste RNK: ribosomalna, najdaljša, ki tvori številne zanke, najkrajša informacijska (matrica) in transportna. Ribosomska RNA je lokalizirana v ribosomih endoplazmatskega retikuluma in predstavlja 85% celotne RNA celice.
Messenger RNA je strukturiran kot list detelje. Njegova količina je 5% celotne RNA v celici. Sintetizira se v nukleolih. Sestavljanje poteka v kromosomih v medfaznem obdobju. Njegova glavna funkcija je prenos informacij iz DNK v ribosome, kjer pride do sinteze beljakovin.
Kot je zdaj ugotovljeno, je prometna RNA cela družina spojin, ki so povezane v strukturi in biološkem delovanju. Vsaka živa celica po približni oceni vsebuje 40-50 posameznih RNK \u200b\u200bza prevoz, njihovo skupno število v naravi pa je, upoštevajoč razlike v vrstah, ogromno. (Acad. W. Engelhardt). Imenujejo jih transport, ker se njihove molekule ukvarjajo s prevoznimi storitvami za znotrajcelični proces sinteze beljakovin. V kombinaciji z brezplačnimi aminokislinami jih dostavljajo ribosomom v verigi gradbenih beljakovin. To so najmanjše molekule RNA, v povprečju 80 nukleotidov. Lokalizirani v citoplazemskem matriksu in tvorijo približno 10% celične RNA
RNA vsebuje štiri dušikove baze, vendar za razliko od DNK molekula RNA vsebuje uracil namesto timina.
Struktura kromozeKromosome so prvič odkrili konec 19. stoletja klasika citologije Fleming in Strasburger (1882, 1884) ter ruski raziskovalec celic I.D. Čistjakov jih je odkril leta 1874.
Glavni strukturni element kromoze je jedro. Imajo različne oblike. To so ravne ali ukrivljene palice, ovalna telesa, kroglice, katerih velikosti se razlikujejo.
Glede na lokacijo centromerja obstajajo ravni, enaki kraki in neenaki kromosomi. Notranja struktura kromoze je prikazana na Sl. 15, 16. Upoštevati je treba, da je deoksiribonukleoprotein kromosomski monomer.
V kromosomu je deoksiribonukleoproteinov 90-92%, od tega 45% DNK in 55% beljakovin (histon). RNA (informativna) je v majhni količini predstavljena tudi v kromosomu.
Kromosom ima tudi jasno izraženo prečno strukturo - prisotnost odebeljenih območij - diskov, ki segajo že leta 1909. so se imenovali geni. Ta izraz je predlagal danski znanstvenik Johannsen. Leta 1911 je ameriški znanstvenik Morgan dokazal, da so geni glavne dedne enote in se v kromosomih razporejajo v linearnem vrstnem redu, zato ima kromosom kvalitativno različna področja. Leta 1934 je ameriški znanstvenik Pinter dokazal prekinitev morfološke strukture kromos in prisotnost diskov v kromosomih, diski pa so kraji, kjer se nabira DNK. To je bil začetek ustvarjanja zemljevidov kromosomov, ki so navajali lokacijo (lokus) gena, ki določa določeno značilnost organizma. Gen je odsek dvojne vijačnice DNA, ki vsebuje podatke o strukturi enega samega proteina. To je odsek molekule DNA, ki določa sintezo ene molekule proteina. DNK ni neposredno vključena v sintezo beljakovin. Vsebuje in hrani samo podatke o strukturi beljakovin.
Struktura DNK, sestavljena iz več tisoč zaporedno lociranih 4 nukleotidov, je označena za dednost.
Koda dednosti Sinteza beljakovinPrvo sporočilo o kode DNA je ameriški biokemik Nirenberg leta 1961 v Moskvi na Mednarodnem biokemičnem kongresu. Bistvo kode DNK je naslednje. Vsaka aminokislina ustreza delu verige DNA treh sosednjih nukleotidov (tripleta). Torej, na primer, mesto, sestavljeno iz T-T-T (trojica treh nukleotidov, ki vsebujejo timine), ustreza aminokislini lizin, tripletu A (adenin) - C (citozin) - A (adenin) - cisteinu itd. Predpostavimo, da je gen predstavljen z verigo nukleotidov, razporejenih v naslednjem vrstnem redu: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G. Ko bomo to vrstico razdelili na trojčke, bomo takoj razvozlali, katere aminokisline in v kakšnem vrstnem redu se bodo nahajale v sintetiziranem proteinu.
Število možnih kombinacij 4 razpoložljivih treh nukleotidov je 4 × 64. Na podlagi teh razmerij je število različnih trojčkov več kot dovolj za zagotavljanje informacij o sintezi številnih beljakovin, ki določajo tako strukturo kot tudi funkcije telesa. Za sintezo beljakovin se ribosomom pošlje natančna kopija teh informacij v obliki messenger RNA. Poleg i-RNA sodeluje pri dekodiranju in sintezi veliko število molekul različnih transportnih ribonukleinskih kislin (t-RNA), ribosomi in številni encimi. Vsaka od 20 aminokislin se veže na T-RNA - molekulo na molekulo. Vsaka od 20 aminokislin ima svojo t-RNA. T-RNA ima kemične skupine, ki lahko "prepoznajo" njegovo aminokislino in jo izberejo med razpoložljivimi aminokislinami. To se zgodi s pomočjo posebnih encimov. Potem ko je prepoznala njegovo aminokislino, t-RNA stopi v povezavo z njo. Na začetku verige (molekule) i-RNA je pritrjen ribosom, ki se giblje vzdolž i-RNA med seboj v polipeptidni verigi poveže točno tiste aminokisline, katerih vrstni red je šifriran z nukleotidnim zaporedjem te I-RNA. Kot rezultat tega nastane beljakovinska molekula, katere sestava je kodirana v enem od genov.
Nucleoli- sestavni del jedra. To so sferična telesa. So zelo spremenljivi, spremenijo obliko in strukturo, se pojavijo in izginejo. Obstajata eno, dve. Za vsako rastlino obstaja določeno število. Nukleoli izginejo, ko se celica pripravi na delitev in se nato ponovno pojavijo; zdi se, da sodelujejo pri sintezi ribonukleinskih kislin. Če je nukleolus uničen s fokusiranim žarkom rentgenskih žarkov ali ultravijoličnih žarkov, potem delitev celic zatremo.
Vloga jedra v življenju celice. Jedro služi kot kontrolno središče celice - usmerja celično aktivnost in vsebuje prenašalce dednosti (gene), ki določajo značilnosti danega organizma. Vlogo jedra lahko razkrijemo, če ga odstranimo iz celice z mikrokirurškimi tehnikami in opazimo posledice tega. Gemmerling je na enocelični zeleni algi Acetobularia izvedel številne poskuse, ki so dokazali pomembno vlogo pri uravnavanju rasti celic. Ta morska alga doseže višino 5 cm, izgleda kot goba, ima videz "korenin" in "nog". Na vrhu se konča z velikim "pokrovčkom" v obliki diska. Celica te alge ima eno jedro, ki se nahaja v bazalnem delu celice.
Hammerling je ugotovil, da če odrežemo steblo, spodnji del še naprej živi in \u200b\u200bpo operaciji popolnoma regenerira pokrovko. Zgornji del, ki je brez jedra, nekaj časa preživi, \u200b\u200bvendar na koncu umre, spodnjega dela pa ne more obnoviti. Zato je jedro acetobularije potrebno za presnovne reakcije, na katerih temelji rast.
Jedro prispeva k nastanku celične membrane. To lahko ponazorimo s poskusi z algama Voucheria in Spyrogyra. S sprostitvijo vsebine celic iz rezanih filamentov v vodo lahko dobimo grudice citoplazme z enim, z več jedri in brez jeder. V prvih dveh primerih se je celična membrana oblikovala normalno. Ker ni jedra, ni nastala lupina.
V poskusih I. I. Gerasimova (1890) s spirogiro so ugotovili, da celice z dvojnim jedrom podvojijo dolžino in debelino kloroplasta. V celicah brez jedra se proces fotosinteze nadaljuje, nastane asimilativni škrob, hkrati pa proces njegove hidrolize izumre, kar je razloženo z odsotnostjo hidroliznih encimov, ki bi jih lahko v ribosomih sintetizirali le glede na podatke jedrske DNK. Življenje protoplasta brez jedra je nepopolno in kratkotrajno. V poskusih I.I. Gerasimove celice Spirogyra brez jedra so živele 42 dni in umrle. Ena najpomembnejših funkcij jedra je oskrba citoplazme z ribonukleinsko kislino, ki je potrebna za sintezo beljakovin v celici. Odstranjevanje jedra iz celice vodi v postopno zmanjšanje vsebnosti RNA v citoplazmi in upočasnitev sinteze beljakovin v njej.
Najpomembnejša vloga jedra pri prenosu znakov s celice na celico, z organizma na organizem in to izvaja v procesu delitve jedra in celice kot celote.
Delitev celic.Celice se množijo z delitvijo. V tem primeru se iz ene celice tvorita dve hčerinski celici z enakim naborom dednega materiala, ki ga vsebuje kromosom kot matična celica. V somatskih celicah so kromosomi predstavljeni z dvema, tako imenovanima homolognima kromosomoma, v katera so vdelani alelni geni (nosilci nasprotnih lastnosti, na primer bela in rdeča barva cvetnih listov itd.), Lastnosti dveh starševskih parov. V zvezi s tem je v somatskih celicah rastlinskega telesa vedno podvojen nabor kromoz, označen z 2n. Kromosomi imajo izrazito osebnost. Količina in kakovost kromoze sta značilni za vsako vrsto. Torej, v jagodnih celicah je diploidni sklop kromoze 14, (2n), jabolko -34, jeruzalem artičoka - 102 itd.
Mitoza (kariokineza) - delitev somatskih celic sta prvič opisala E. Russov (1872) in I. D. Čistjakov (1874). Njegovo bistvo je v tem, da se iz matične celice z delitvijo tvorita dve hčerinski celici z enakim naborom kromoz, ki sta sestavljeni iz interfaze in same mitoze. Z metodo mikroavtoradiografije je bilo ugotovljeno, da je najdaljša in najtežja interfaza - obdobje jedra »počivajočega«, ker v tem obdobju se podvoji jedrski material. Interfaza je razdeljena na tri faze:
Q1 - presintetični (njegovo trajanje je 4-6 ur);
S - sintetična (10-20 ur);
Q2 - post-sintetično (2-5 ur).
Med fazo Q1 potekajo priprave za redukcijo DNK. In v S-fazi pride do redukcije DNA, celica podvoji svojo zalogo DNK. V fazi Q2 se tvorijo encimi in strukture, ki so potrebni za sprožitev mitoze. Tako se v interfazi molekule DNK v kromosomih razdelijo na dva enaka pramena, informacijske RNA pa so sestavljene na njihovi matrici. Slednji nosi podatke o zgradbi specifičnih beljakovin v citoplazmi, v jedru pa vsak od verig DNK dopolni manjkajočo polovico njegove molekule. V tem procesu podvajanja (redukcije) se kaže edinstvena lastnost DNK, ki je sestavljena iz sposobnosti DNK, da se natančno reproducira. Tako oblikovane hčerinske molekule DNK se samodejno pridobijo kot natančne kopije matične molekule, saj med reduplikacijo se na vsako polovico pritrdijo komplementarne (AT; G-C; itd.) podlage iz okolja.
Med fazo mitotske delitve postanejo podvojeni kromosomi opazni. V metafazi so vsi nameščeni v ekvatorialni coni, razporejeni v eni vrsti. Navoji vretena so oblikovani (iz mikrotubul, ki se povezujejo med seboj). Lupina jedra in nukleolus izgineta. Zadebljeni kromosomi se razdelijo po dolžini na dva hčerinska kromosoma. To je bistvo mitoze. Zagotavlja natančno porazdelitev dvojnih molekul DNK med hčerinskimi celicami. Tako zagotavlja tudi prenos dednih informacij, kodiranih v DNK.
V anafazi se hčerinski kromosomi začnejo premikati na nasprotne polove. Prvi drobci celične membrane (fragmoblast) se pojavijo v središču.
V telofazi nastajajo jedra v hčerinskih celicah. Vsebina matične celice (organele) se porazdeli med tvorjene hčerinske celice. Celična membrana je v celoti oblikovana. S tem se konča citokineza (slika 17).
Mejoza - oddelek za zmanjšanje je odkril in opisal v 90. letih prejšnjega stoletja V. I. Belyaev. Bistvo delitve je v dejstvu, da se iz somatske celice, ki vsebuje 2n (dvojni, diploidni) sklop kromoz, tvorijo štiri haploidne celice, z "n", pol nabora kromoz. Ta vrsta delitve je zapletena in je sestavljena iz dveh stopenj. Prva je redukcija s kromozo. Dvojni kromosomi se v paru (dva vzporedna homologna kromosoma) nahajajo v ekvatorialni coni. V tem trenutku lahko pride do konjugacije (povezave) kromoze, prečkanja (prečkanja) in posledično do izmenjave mest kromoze. Posledično se nekateri geni očetovskih kromos prenesejo v materinske kromoze in obratno. Posledično se videz obeh kromos ne spremeni, vendar njihova kakovostna sestava postane drugačna. Očetovsko in materinsko dedovanje se prerazporedi in meša.
V anafazi mejoze se homologni kromosomi s pomočjo vretenastih filamentov razhajajo vzdolž polov, na katerih se po kratkem času mirovanja (nitke izginejo, septum med novimi jedri pa ne tvori) pa se začne proces mitoze - metafaza, pri kateri se vsi kromosomi nahajajo v isti ravnini in pride do njihovega vzdolžnega cepitve na hčerinskih kromosomih. Med anafazo mitoze s pomočjo vretena se razhajajo do polov, kjer se tvorijo štiri jedra in posledično štiri haploidne celice. V celicah nekaterih tkiv se med njihovim razvojem pod vplivom nekaterih dejavnikov pojavi nepopolna mitoza in število kromoz v jedrih se podvoji zaradi dejstva, da se na polov ne razhajajo. Kot posledica takšnih kršitev naravne ali umetne narave nastanejo tetraploidni in poliploidni organizmi. S pomočjo mejoze se tvorijo spolne celice - gamete, pa tudi spore, elementi spolnega in aseksualnega razmnoževanja rastlin (slika 18).
Amitoza je neposredna cepitev jedra. Pri amitozi se fisijsko vreteno ne tvori in jedrska membrana se ne razgradi, kot pri mitozi. Pred tem je amitoza veljala za primitivno obliko delitve. Zdaj je bilo ugotovljeno, da je povezano z razgradnjo telesa. Gre za poenostavljeno različico kompleksnejše jedrske cepitve. Amitoza se pojavi v celicah in tkivih nukleusa, endosperma, parenhima gomoljev, listnih peteljk itd.
Mitohondrije je spiralna, zaobljena, podolgovata ali razvejena organela.
Koncept mitohondrije je Benda prvič predlagala leta 1897. Mitohondrije lahko najdemo v živih celicah z uporabo faznega kontrasta in interferenčne mikroskopije v obliki zrn, zrnc ali nitk. To so precej mobilne strukture, ki se lahko premikajo, združujejo med seboj in delijo. Ob obarvanju s posebnimi metodami v odmrlih celicah pod svetlobno mikroskopijo imajo mitohondrije v obliki majhnih zrn (zrnc), difuzno razporejenih v citoplazmi ali koncentriranih na nekaterih njegovih določenih območjih.
Zaradi uničenja glukoze in maščob v prisotnosti kisika nastaja energija v mitohondrijih, organske snovi pa se pretvorijo v vodo in ogljikov dioksid. Na ta način živalski organizmi dobijo glavno energijo, potrebno za življenje. Energija se nabira v adenozin trifosfatu (ATP), ali bolje rečeno, v njegovih visokoenergijskih vezicah. Funkcija mitohondrijev je tesno povezana z oksidacijo organskih spojin in uporabo energije, ki se sprosti med njihovim razpadom, za sintezo molekul ATP. Zato se mitohondriji pogosto rečejo energetske postaje celice ali organele celičnega dihanja. ATP deluje kot dobavitelj energije s prenosom ene od svojih energijsko bogatih fosfatnih terminalnih skupin na drugo molekulo in v procesu se pretvori v ADP.
Domnevajo, da so bili mitohondriji v evoluciji prokariontni mikroorganizmi, ki so postali simbiotični v telesu starodavne celice. Nato so postali vitalni, kar je bilo povezano s povečanjem vsebnosti kisika v Zemljini atmosferi. Po eni strani so mitohondriji odstranili odvečni kisik, ki je strupen za celico, na drugi strani pa so zagotovili energijo.
Brez mitohondrije celica praktično ne more uporabljati kisika kot snovi, ki zagotavlja energijo, in lahko samo z anaerobnimi procesi napolni svoje energijske potrebe. Tako je kisik strup, vendar je strup življenjsko pomemben za celico, presežek kisika pa je prav tako škodljiv kot njegovo pomanjkanje.
Mitohondriji lahko spremenijo svojo obliko in se preselijo na tista območja celice, kjer je potreba po njih največja. Torej, v kardiomiocitih so mitohondriji nameščeni v bližini miofibrilov, v celicah ledvičnih tubulov v bližini bazalnih invaginacij itd. Celica vsebuje do tisoč mitohondrijev, njihovo število pa je odvisno od aktivnosti celice.
Mitohondrije imajo povprečno prečno velikost 0,5 ... 3 mikrona. Glede na velikost ločimo majhne, \u200b\u200bsrednje, velike in velikanske mitohondrije (tvorijo razvejeno mrežo - mitohondrijski retikulum). Velikost in število mitohondrijev sta tesno povezana z aktivnostjo celice in njeno porabo energije. So izjemno spremenljive in glede na aktivnost celice lahko vsebnost kisika, hormonski vplivi nabreknejo, spremenijo število in strukturo kristov, se razlikujejo po številu, obliki in velikosti ter encimski aktivnosti.
Volumetrična gostota mitohondrijev, stopnja razvoja njihove notranje površine in drugi kazalniki so odvisni od energijskih potreb celice. V limfocitih je le nekaj mitohondrijev, v jetrnih celicah pa 2 ... 3 tisoč.
Mitohondrije sestavljajo matrica, notranja membrana, perimitohondrijski prostor in zunanja membrana. Zunanja mitohondrijska membrana loči organelo od hijaloplazme. Običajno ima gladke konture in je zaprt, tako da je membranska vrečka.
Zunanja membrana je ločena od notranje membrane s perimitohondrijskim prostorom, širokim približno 10 ... 20 nm. Notranja mitohondrijska membrana omejuje dejansko notranjo vsebnost mitohondrijev - matriks. Notranja membrana tvori številne izrastke v mitohondrijah, ki so videti kot ploščati grebeni ali križe.
Po obliki imajo kriste lahko obliko plošč (trabekularne) in tubule (multivetikularne na rezu), usmerjene pa so vzdolžno ali prečno glede na mitohondrije.
Vsak mitohondrij je napolnjen z matrico, ki je na elektronskih mikrografih videti gostejša kot okoliška citoplazma. Mitohondrijska matrica je homogena (homogena), včasih drobnozrnata, z različno gostoto elektronov. Razkriva tanke niti približno 2 ... 3 nm in zrnca približno 15 ... 20 nm. Matrični prameni so molekule DNK, majhna zrnca pa mitohondrijski ribosomi. Matrica vsebuje encime, en enoveričen ciklični DNK, mitohondrijske ribosome, veliko Ca 2+ ionov.
Avtonomni sistem sinteze proteinov mitohondrijev predstavlja molekule DNK brez histona. DNK je kratek, obročast (cikličen) in vsebuje 37 genov. Za razliko od jedrske DNK v njej praktično ni nekodirajočih nukleotidnih sekvenc. Značilnosti strukture in organizacije približajo mitohondrijsko DNK DNK bakterijskih celic. Molekule RNK se sintetizirajo na mitohondrijski DNK različni tipi: informacijski, prenos (transport) in ribosomalni. RNA mitohondrijskega sesalca se ne spoji (izrezovanje področij, ki ne prenašajo informacijske obremenitve). Majhna velikost mitohondrijskih molekul DNK ne more določiti sinteze vseh mitohondrijskih beljakovin. Večina mitohondrijskih beljakovin je pod genskim nadzorom celičnega jedra in se sintetizira v citoplazmi, saj je mitohondrijska DNK slabo izražena in lahko zagotovi tvorbo le dela encimov oksidativne fosforilacijske verige. Mitohondrijska DNK ne kodira več kot deset beljakovin, ki so lokalizirane v membranah in so strukturni proteini, odgovorni za pravilno integracijo posameznih funkcionalnih beljakovinskih kompleksov mitohondrijskih membran. Sintetizirajo se tudi beljakovine, ki izvajajo transportne funkcije. Tak sistem sinteze beljakovin ne zagotavlja vseh funkcij mitohondrijev, zato je mitohondrijska avtonomija omejena in relativna.
Pri sesalcih se med oploditvijo mitohondriji prenašajo samo skozi jajčece, semenčice pa v nov organizem vnašajo jedrsko DNK.
V matriksu mitohondrijev nastajajo ribosomi, ki se razlikujejo od ribosomov citoplazme. Vključeni so v sintezo številnih mitohondrijskih beljakovin, ki jih jedro ne kodira. Mitohondrijski ribosomi imajo sedimentacijsko število 60 (v nasprotju s citoplazemskimi ribosomi z usedalnim številom 80). Številčno usedanje je hitrost usedanja za centrifugiranje in ultracentrifugiranje. Mitohondrijski ribosomi so po strukturi blizu ribosomov prokariotskih organizmov, vendar so manjši in se razlikujejo po občutljivosti na nekatere antibiotike (kloramfenikol, tetraciklin itd.).
Notranja mitohondrijska membrana ima visoko stopnjo selektivnosti pri transportu snovi. Na njegovo notranjo površino so pritrjeni encimi oksidativne fosforilacijske verige, elektronski transportni proteini, transportni sistemi ATP, ADP, piruvat. učinkovitost katalitičnih procesov.
Elektronska mikroskopija razkriva delce gob, ki štrlijo v lumen matriksa. Imajo sintetično ATP (tvori ATP iz ADP). Elektroni se prevažajo po dihalni verigi, ki se nahaja v notranji membrani, ki vsebuje štiri velike encimske komplekse (citokromi). Ko elektroni prehajajo skozi dihalno verigo, se vodikovi ioni izčrpajo iz matrike v perimitohondrijski prostor, kar zagotavlja nastanek protonskega gradienta (črpalke). Energija tega gradienta (razlike v koncentraciji snovi in \u200b\u200btvorba membranskega potenciala) se uporablja za sintezo ATP in transport metabolitov in anorganskih ionov. Nosilni proteini, ki jih vsebuje notranja membrana, preko nje prenašajo organske fosfate, ATP, ADP, aminokisline, maščobne kisline, tri- in dikarboksilne kisline.
Zunanja membrana mitohondrijev je bolj prepustna za snovi z nizko molekulsko maso, saj vsebuje veliko hidrofilnih beljakovinskih kanalov. Na zunanji membrani so nameščeni specifični receptorski kompleksi, skozi katere se proteini iz matrike prenašajo v perimitohondrijski prostor.
Po svoji kemični sestavi in \u200b\u200blastnostih je zunanja membrana blizu drugih medceličnih membran in plazmolemme. Vsebuje encime, ki presnavljajo maščobe, aktivirajo (katalizirajo) pretvorbo aminov, amin oksidazo. Če encimi zunanje membrane ostanejo aktivni, je to pokazatelj funkcionalne varnosti mitohondrijev.
V mitohondrijah obstajata dve avtonomni poddružini. Če se perimitohondrijski prostor ali zunanja komora mitohondrijev (zunanji pododdelek) oblikuje zaradi prodora beljakovinskih kompleksov hijaloplazme, potem se zaradi sintetične aktivnosti delno oblikuje notranji pododdelek (mitohondrijski matriks). mitohondrijska DNA... Notranji pododdelek (matrica) vsebuje DNK, RNA in ribosome. Ima visoko raven Ca 2+ ionov v primerjavi s hiialoplazmo. Vodikovi ioni se kopičijo v zunanjem poddelku. Encimatska aktivnost zunanjih in notranjih delcev ter sestava beljakovin sta zelo različni. Notranji pododdelek ima večjo gostoto elektronov kot zunanji.
Specifični markerji mitohondrije so encimi citokrom oksidaza in sukcinatna dehidrogenaza, katerih prepoznavanje omogoča kvantitativno karakterizacijo energijskih procesov v mitohondrijih.
Glavna funkcija mitohondrijev - sinteza ATP. Sprva se v hijaloplazmi sladkorji (glukoza) uničijo do mlečne in piruvične kisline (piruvat) s hkratno sintezo majhne količine ATP. Kot posledica glikolize ene molekule glukoze uporabljamo dve molekuli ATP in nastanejo štiri. Tako le dve molekuli ATP sestavljata pozitivno ravnovesje. Ti procesi potekajo brez kisika (anaerobna glikoliza).
Vsi nadaljnji koraki pri proizvodnji energije se pojavljajo v procesu aerobne oksidacije, kar zagotavlja sintezo velikih količin ATP. V tem primeru se organske snovi uničijo do CO 2 in vode. Oksidacijo spremlja prenos protonov na njihove sprejemnike. Te reakcije izvajajo številni encimi cikla trikarboksilne kisline, ki se nahajajo v matriksu mitohondrijev.
Sistemi prenosa elektronov in s tem povezano fosforilacija ADP (oksidativno fosforilacijo) so vgrajeni v membrane kriz. V tem primeru se elektroni prenašajo iz enega proteina akceptorjev elektronov v drugega in se na koncu vežejo s kisikom, zaradi česar nastane voda. Hkrati se del energije, ki se sprosti med takšno oksidacijo v verigi prenosa elektronov, shrani v obliki visokoenergijske vezi med fosforilacijo ADP, kar vodi do tvorbe velikega števila molekul ATP, glavnega medceličnega ekvivalenta energije. Na membranah mitohondrijskih kriz se proces oksidativne fosforilacije zgodi s pomočjo beljakovin oksidacijske verige in fosforilacijskega encima ADP sintetaze ATP, ki se nahaja tukaj. Kot posledica oksidativne fosforilacije nastane 36 molekul ATP iz ene molekule glukoze.
Za nekatere hormone in snovi obstajajo specializirani (afinitetni) receptorji na mitohondrijih. Trijodtironin običajno pospeši sintetično aktivnost mitohondrijev. Interlevkin-1 in visoke koncentracije trijodotironina odklopijo verige oksidacijske fosforilacije, povzročijo otekanje mitohondrijev, kar spremlja povečanje proizvodnje toplotne energije.
Novi mitohondriji nastanejo z delitvijo, zožitvijo ali popki. V slednjem primeru se oblikuje protomitohondrion, ki se postopoma povečuje v velikosti.
Protomitohondrija je majhna organela z zunanjo in notranjo membrano. Notranja membrana nima ali vsebuje nerazvitih kriz. Za organelo je značilen: nizka stopnja aerobna fosforilacija. Z nastankom zožitve se vsebnost mitohondrijev porazdeli med dva nova dokaj velika organela. Pri kateri koli metodi razmnoževanja ima vsak novo nastali mitohondrij svoj genom.
Stare mitohondrije uničijo z avtolizo (samo prebavo s celico z uporabo lizosomov), da tvorijo avtolizosome. Iz avtolizosoma nastane preostalo telo. S popolno prebavo je vsebnost preostalega telesa, sestavljena iz nizke molekulske mase organska snov, se izloča z eksocitozo. Z nepopolno prebavo se lahko v celici naberejo ostanki mitohondrijev v obliki slojevitih teles ali granul z nipofuscinom. V nekaterih mitohondrijah se kopičijo netopne kalcijeve soli s tvorbo kristalov - kalcifikacij. Kopičenje produktov degeneracije mitohondrijev lahko privede do degeneracije celic.
Struktura in delovanje mitohondrijev je zapleteno vprašanje. Prisotnost organele je značilna za skoraj vse jedrske organizme, tako za avtotrofe (rastline, sposobne fotosinteze) kot za heterotrofe, ki so skoraj vse živali, nekatere rastline in glive.
Glavni namen mitohondrijev je oksidacija organskih snovi in \u200b\u200bnadaljnja uporaba energije, ki se sprosti kot rezultat tega procesa. Zaradi tega imajo organele tudi drugo (neuradno) ime - energetske postaje celice. Včasih jih imenujejo "katabolični plastidi".
Kaj so mitohondriji
Izraz je grškega izvora. V prevodu ta beseda pomeni "nit" (mitos), "zrno" (hondrion). Mitohondrije so trajne organele, ki so velikega pomena za normalno delovanje celic in omogočajo obstoj celotnega organizma.

„Postaje“ imajo posebno notranjo strukturo, ki se spreminja glede na funkcionalno stanje mitohondrijev. Njihova oblika je lahko dve vrsti - ovalna ali podolgovata. Slednji ima pogosto razvejan videz. Število organelov v eni celici se giblje od 150 do 1500.
Poseben primer so spolne celice. Spermatozoji vsebujejo le eno spiralno organelo, medtem ko ženske gamete vsebujejo sto tisoč mitohondrijev. Organele v celici niso pritrjene na enem mestu, ampak se lahko premikajo vzdolž citoplazme, se med seboj kombinirajo. Njihova velikost je 0,5 mikrona, dolžina lahko doseže 60 mikronov, najmanjši indikator pa 7 mikronov.

Določitev velikosti ene same elektrarne ni lahka naloga. Dejstvo je, da ob gledanju z elektronskim mikroskopom v delček pade le del organele. Tako se zgodi, da ima spiralni mitohondrij več odsekov, ki jih lahko jemljemo kot ločene, neodvisne strukture.
Le tridimenzionalna slika vam bo omogočila, da ugotovite natančno celično zgradbo in razumete, ali govorimo o 2-5 ločenih organelov ali o eni mitohondriji, ki ima zapleteno obliko.
Značilnosti strukture
Mitohondrijska membrana je sestavljena iz dveh plasti: zunanje in notranje. Slednji vključuje različne izrastke in nabore, ki imajo listnato in cevasto obliko.
Vsaka membrana ima posebno kemično sestavo, določeno količino določenih encimov in določen namen. Zunanja lupina je ločena od notranje lupine z medoslobnim prostorom debeline 10-20 nm.
Struktura organele je na sliki zelo jasno videti z napisi.

Diagram strukture mitohondrijev
Če pogledate strukturni diagram, lahko naredite naslednji opis. Viskozni prostor znotraj mitohondrije imenujemo matrica. Njegova sestava ustvarja ugodno okolje za potrebne kemične procese v njem. Vsebuje mikroskopske granule, ki spodbujajo reakcije in biokemične procese (na primer kopičijo ione glikogena in drugih snovi).
Matrica vsebuje DNK, koencime, ribosome, t-RNA, anorganske ione. ATP sintaza in citokromi se nahajajo na površini notranje plasti lupine. Encimi prispevajo k procesom, kot so Krebsov cikel (CTK), oksidativna fosforilacija itd.
Tako glavno nalogo organoida opravlja tako matrica kot notranja stran lupine.
Mitohondrijske funkcije
Namen "elektrarn" lahko označimo z dvema glavnima nalogama:
- proizvodnja energije: v njih potekajo oksidativni procesi, ki jim sledi sproščanje molekul ATP;
- shranjevanje genetskih informacij;
- sodelovanje v sintezi hormonov, aminokislin in drugih struktur.
Postopek oksidacije in pridobivanja energije poteka v več fazah:


Shematski prikaz sinteze ATP
Omeniti velja: kot posledica Krebsovega cikla (cikla citronske kisline) se molekule ATP ne tvorijo, molekule se oksidirajo in sprosti ogljikov dioksid. To je vmesni korak med glikolizo in elektronsko transportno verigo.
Tabela "Funkcije in struktura mitohondrijev"

Kaj določa število mitohondrijev v celici
Prevladujoče število organelov se nabira v bližini tistih delov celice, kjer so potrebe po energijskih virih. Zlasti se na območju miofibrilov nabere veliko število organelov, ki so del mišičnih celic, ki zagotavljajo njihovo krčenje.

V moških zarodnih celicah so strukture lokalizirane okoli osi flagelluma - domneva se, da je potreba po ATP posledica nenehnega gibanja repa gamete. Razporeditev mitohondrijev v protozojih je videti popolnoma enaka, pri čemer za gibanje uporabljajo posebne cilije - organele se kopičijo pod membrano na njihovem dnu.
Kar zadeva živčne celice, opazimo lokalizacijo mitohondrijev blizu sinaps, skozi katere se prenašajo signali živčnega sistema. V celicah, ki sintetizirajo beljakovine, se organele kopičijo v conah ergastoplazme - oskrbujejo z energijo, ki zagotavlja ta proces.
Kdo je odkril mitohondrije
Celicna struktura je svoje ime dobila v letih 1897-1898 po K. Brandu. Povezavo med procesi celičnega dihanja in mitohondrijev je leta 1920 dokazal Otto Wagburg.
Zaključek
Mitohondrije so najpomembnejši sestavni del žive celice, saj delujejo kot energetska postaja, ki proizvaja molekule ATP in s tem zagotavlja procese celičnega življenja.
Delo mitohondrijev temelji na oksidaciji organskih spojin, kar ima za posledico ustvarjanje energijskega potenciala.
Zunanja membrana
Notranja membrana
Matrica m-na, matrica, crista... ima gladke konture, ne tvori izrastkov ali gub. Sestavlja približno 7% površine vseh celičnih membran. Njegova debelina je približno 7 nm, ni povezana z nobenimi drugimi membranami citoplazme in je zaprta vase, tako da je membranska vrečka. Zunanja membrana je ločena od notranje medmestni prostor približno 10-20 nm širok. Notranja membrana (debela približno 7 nm) omejuje dejansko notranjo vsebnost mitohondrijev,
njegova matrica ali mitoplazma. Značilna značilnost notranje membrane mitohondrijev je njihova sposobnost tvorjenja številnih invagin v mitohondrije. Takšne invazije imajo najpogosteje v obliki ravnih grebenov ali križ. Razdalja med membranami v kristi je približno 10-20 nm. Kriste se pogosto lahko vejo ali tvorijo prstne procese, se upognejo in nimajo izrazite orientacije. V protozojih, enoceličnih algah, v nekaterih celicah višjih rastlin in živali so izrastki notranje membrane videti kot cevi (tubular cristae).
Mitohondrijska matrica ima fino zrnato homogeno strukturo, včasih se v njej razkrijejo tanki nitki, zbrani v kroglico (približno 2-3 nm) in zrnca približno 15-20 nm. Zdaj je postalo znano, da so nitki mitohondrijskega matriksa molekule DNA v mitohondrijskem nukleoidu, majhna zrnca pa mitohondrijski ribosomi.
Mitohondrijske funkcije
1. Sinteza ATP se pojavi v mitohondrijih (glejte oksidativno fosforilacijo)
PH medmembranskega prostora ~ 4, pH matrice ~ 8 | vsebnost beljakovin v m: 67% - matrica, 21% - zunaj m-on, 6% - znotraj m-on in 6% - v inter-m-on
Handrioma - enoten sistem mitohondrijev
zunanja plast: porozne pore omogočajo prehod do 5 kD | notranji mn: \u200b\u200bkardiolipin - naredi neprepustne za ione |
medmnožna tvorba: encimske skupine fosforilirajo nukleotide in sladkorje nukleotide
notranji m-on:
matrika: presnovni encimi - oksidacija lipidov, oksidacija ogljikovih hidratov, cikel trikarboksilne kisline, Krebsov cikel
Izvor bakterij: ameba Pelomyxa palustris ne vsebuje m. Evkariontov, živi v simbiozi z aerobnimi bakterijami | lastna DNK | procesi, podobni bakterijam
Mitohondrijska DNA
Oddelek miohondrijev
podvojeno
v interfazi | podvajanje ni povezano s S-fazo | med celičnim ciklom se mitoh enkrat razdeli na dva, tvori zožitev, zoženje najprej na notranjem m-ne | ~ 16,5 kb | krožno, kodira 2 rRNA, 22 tRNA in 13 proteinov |
transport beljakovin: signalni peptid | amfifilni curl | receptor za prepoznavanje mitohondrijev |
Oksidativna fosforilacija
Transportna veriga elektronov
ATP sintaza
v jetrnih celicah m živi ~ 20 dni delitev mitohondrijev s tvorbo zožitve
16569 bp \u003d 13 proteinov, 22tRNA, 2pRNA | gladek zunanji mineral (porine - prepustnost beljakovin do 10 kDa) zložen notranji (cristae) mineral (75% beljakovin: transportni proteini, transporterji, f-vi, sestavine dihalne verige in ATP sintaza, kardiolipin) matriks ( obogatena s f-tami citratnega cikla) \u200b\u200bmedmnožna proizvodnja 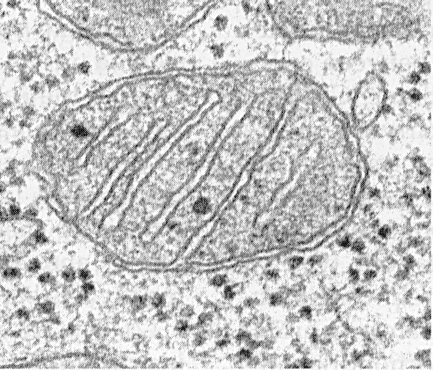
Mitohondriji ali hondriosomi (iz grške mitos - nit, hondrion - zrno, soma - malo telo) so zrnate ali nitaste organele, prisotne v citoplazmi protozojev, rastlin in živali. Mitohondrije lahko opazimo v živih celicah, saj imajo dokaj visoko gostoto. V živih celicah se mitohondriji lahko premikajo, premikajo, združujejo med seboj.
Imej različni tipi velikost mitohondrijev je zelo spremenljiva, prav tako kot je njihova oblika spremenljiva (slika 199). Kljub temu je v večini celic debelina teh struktur relativno konstantna (približno 0,5 mikrona), dolžina pa niha in v nitastih oblikah doseže 7-60 mikronov.
Preučevanje velikosti in števila mitohondrijev ni tako preprosta zadeva. To je posledica dejstva, da velikost in število mitohondrij, opaženih na ultra tankih odsekih, ne ustrezata resničnosti.
Običajni izračuni kažejo, da je približno 200 mitohondrijev na jetrno celico. To predstavlja več kot 20% celotne citoplazme in približno 30-35% celotne beljakovine v celici. Površina vseh mitohondrij jetrne celice je 4-5 krat večja od površine njene plazemske membrane. Največ mitohondrijev je v oocitih (približno 300.000) in v velikanskem kaosu Amee Chaos (do 500.000).
V celicah zelenih rastlin je število mitohondrij manjše kot v živalskih celicah, saj lahko kloroplasti opravljajo del svojih funkcij.
Lokalizacija mitohondrijev v celicah je različna. Običajno se mitohondriji kopičijo v bližini tistih delov citoplazme, kjer obstaja potreba po ATP, ki nastane v mitohondrijah. Torej, v skeletnih mišicah se mitohondrije nahajajo v bližini miofibrilov. Pri spermatozoidih mitohondriji tvorijo spiralno ovojnico okoli osi flagelluma; Verjetno je to posledica potrebe po uporabi ATP za premikanje repa sperme. Podobno se v protozojih in v drugih ciliiliranih celicah mitohondriji lokalizirajo neposredno pod celično membrano na dnu cilijev, za katere je potreben ATP. V aksonih živčnih celic se mitohondrije nahajajo v bližini sinaps, kjer poteka proces prenosa živčnega impulza. V sekretornih celicah, ki sintetizirajo velike količine beljakovin, so mitohondriji tesno povezani z območji ergastoplazme; verjetno dobavljajo ATP za aktiviranje aminokislin in sintezo beljakovin na ribosomih.
Ultrastruktura mitohondrijev.
Mitohondriji imajo ne glede na njihovo velikost ali obliko univerzalno strukturo, njihova ultrastruktura je enotna. Mitohondrije omejujeta dve membrani (slika 205). Zunanja mitohondrijska membrana jo loči od hijaloplazme, ima gladke konture, ne tvori izrastkov ali gub, debelina je približno 7 nm. Sestavlja približno 7% površine vseh celičnih membran. Membrana ni povezana z nobenimi drugimi membranami citoplazme, je zaprta vase in je membranska vreča. Zunanja membrana je od notranje membrane ločena z medemembranskim prostorom, širine približno 10-20 nm. Notranja membrana (debela približno 7 nm) omejuje dejansko notranjo vsebnost mitohondrijev, njegove matrice ali mitoplazme. Notranja mitohondrijska membrana tvori številne invazije v mitohondrije. Takšne invazije imajo najpogosteje v obliki ravnih grebenov ali križ.
Skupna površina notranje mitohondrijske membrane v jetrni celici je približno tretjina površine vseh celičnih membran. Mitohondrije srčnih mišičnih celic vsebujejo trikrat več kriz kot jetrne mitohondrije, kar odraža razlike v funkcionalnih obremenitvah mitohondrij različnih celic. Razdalja med membranami v kristi je približno 10-20 nm.
Mitohondrijske kriste, ki segajo od notranje membrane in segajo proti matriksu, mitohondrijske votline ne blokirajo v celoti in ne motijo \u200b\u200bkontinuitete polnjenja matrike.
Usmerjenost kristov glede na dolgo os mitohondrijev je pri različnih celicah različna. Usmerjenost je lahko pravokotna (jetrne celice, ledvične celice) kristov; v srčni mišici je vzdolžna razporeditev kriz. Kristali se lahko vejo ali tvorijo prstne procese, se upognejo in nimajo izrazite orientacije (sl. 208). V protozojih, enoceličnih algah, v nekaterih celicah višjih rastlin in živali so izrastki notranje membrane videti kot cevi (tubular cristae).
Mitohondrijski matriks ima fino zrnato homogeno strukturo, pri kateri se molekule DNK odkrijejo v obliki tankih nitk, zbranih v kroglico (približno 2-3 nm), in mitohondrijski ribosomi v obliki granul velikosti približno 15-20 nm. Depoziti magnezijevih in kalcijevih soli v matriksu tvorijo velika (20-40 nm) gosta zrna.
Mitohondrijske funkcije.
Mitohondriji izvajajo sintezo ATP, ki nastane kot posledica procesov oksidacije organskih substratov in fosforilacije ADP.
Začetne stopnje oksidacije ogljikovih hidratov se imenujejo anaerobna oksidacija ali glikoliza pojavljajo se v hijaloplazmi in ne potrebujejo kisika. Heksoze in najprej glukoza služijo kot oksidacijski substrat za proizvodnjo anaerobne energije; nekatere bakterije imajo sposobnost pridobivanja energije z oksidacijo pentoz, maščobnih kislin ali aminokislin.
V glukozi je količina potencialne energije, ki jo vsebujejo vezi med atomoma C, H in O, približno 680 kcal na 1 mol (to je 180 g glukoze).
V živi celici se ta ogromna količina energije sprosti v obliki stopenjskega procesa, ki ga nadzorujejo številni oksidativni encimi, in ni povezana s prehodom energije kemijske vezi v toploto, kot pri zgorevanju, ampak prehaja v makroenergetsko vez v molekulah ATP, ki se sintetizirajo ob uporabi sproščene energije iz ADP in fosfat.
Trioze, ki nastanejo kot posledica glikolize, in predvsem pirvična kislina v mitohondrijih, sodelujejo pri nadaljnji oksidaciji. V tem primeru se uporablja energija cepitve vseh kemičnih vezi, kar vodi do sproščanja CO 2, do porabe kisika in sinteze velike količine ATP. Ti procesi so povezani z oksidacijskim ciklom trikarboksilnih kislin in z dihalno transportno verigo elektronov, kjer se ADP fosforilira in se sintetizira celično "gorivo", molekule ATP (slika 209).
V ciklu trikarboksilne kisline (cikel Krebs ali cikel citronske kisline) piruvat, ki nastane kot posledica glikolize, najprej izgubi molekulo CO in se oksidira do acetata (bikarbonska spojina), kombinira s koencimom A. Nato acetil koencim A, združi z oksalacetatom (štiri ogljikova spojina), tvori šest-ogljikov citrat (citronska kislina). Nato sledi cikel oksidacije te šest-ogljikove spojine do štiri ogljikovega oksalacetata, ki se spet veže z acetilnim koencimom A, nato pa se cikel ponovi. Med to oksidacijo se sprostijo dve molekuli CO 2, elektroni, ki se sprostijo med oksidacijo, pa se prenesejo v akceptorske molekule koencimov (NAD-nikotinamid adenin dinukleotid), ki jih nadalje vključijo v verigo prenosa elektronov. V ciklu trikarboksilne kisline torej ni same sinteze ATP, ampak pride do oksidacije molekul, prenosa elektronov do akceptorjev in sproščanja CO 2. Vsi zgoraj opisani dogodki v mitohondriji se odvijajo v njihovi matrici.
Oksidacija začetnega substrata vodi do sproščanja CO 2 in vode, vendar se toplotna energija ne sprošča, kot pri zgorevanju, ampak nastajajo molekule ATP. Sintetizira jih druga skupina beljakovin, ki niso neposredno povezane z oksidacijo. V notranjih mitohondrijskih membranah na površini membran, ki gledajo v matrico, so veliki proteinski kompleksi, encimi in sinteza ATP. V elektronskem mikroskopu jih vidimo v obliki tako imenovanih "gobjih" teles, ki vse obložijo površino membran in gledajo v matrico. Biki imajo nogo in glavo s premerom 8-9 nm. Posledično so encimi tako oksidativne verige kot encimi sinteze ATP lokalizirani v notranjih membranah mitohondrijev (slika 201b).
Dihalna veriga je glavni sistem pretvorbe energije v mitohondrijih. Tukaj pride do konstantne oksidacije in redukcije elementov dihalne verige, zaradi česar se energija sprosti v majhnih delih. Zaradi te energije nastane ATP na treh točkah verige iz ADP in fosfata. Zato pravijo, da je oksidacija (prenos elektrona) povezana s fosforilacijo (ADP + Fn → ATP, to je, da pride do procesa oksidacijske fosforilacije.
Energija, ki se sprosti med transportom elektronov, se shrani v obliki gradienta protonov na membrani. Izkazalo se je, da med prenosom elektronov v mitohondrijski membrani vsak kompleks dihalne verige usmeri brezplačno oksidacijsko energijo za premikanje protonov (pozitivnih nabojev) skozi membrano, iz matrike v medemembranski prostor, kar privede do nastanka potencialne razlike na membrani: pozitivni naboji prevladujejo v medmembranskem prostoru in negativno - iz mitohondrijske matrice. Ko dosežemo razliko potenciala (220 mV), beljakovinski kompleks sintetaze ATP začne prenašati protone nazaj v matrico, medtem ko eno obliko energije pretvori v drugo: tvori ATP iz ADP in anorganskega fosfata. Tako se oksidativni procesi povezujejo s sintetičnim, s fosforilacijo ADP. Medtem ko pride do oksidacije substratov, medtem ko pride do črpanja protonov skozi notranjo mitohondrijsko membrano, poteka povezana ATP sinteza, tj. pride do oksidativne fosforilacije.
Ta dva procesa je mogoče ločiti. V tem primeru se prenos elektronov nadaljuje, prav tako tudi oksidacija substrata, vendar sinteza ATP ne pride. V tem primeru se energija, ki se sprosti med oksidacijo, pretvori v toplotno energijo.
Oksidativna fosforilacija v bakterijah.
V prokariontskih celicah, ki so sposobne oksidativne fosforilacije, so elementi cikla trikarboksilne kisline lokalizirani neposredno v citoplazmi, encimi dihalne verige in fosforilacija pa so povezani s celično membrano, pri čemer njeni izrastki štrlijo v citoplazmo, s tako imenovanimi mezozomi (slika 212). Treba je opozoriti, da so takšni bakterijski mezosomi povezani ne samo s procesi aerobnega dihanja, ampak tudi pri nekaterih vrstah sodelujejo pri celični delitvi, v procesu razdelitve DNK na nove celice, pri nastajanju celične stene itd.
Na plazemski membrani v mezosomih nekaterih bakterij se izvajajo procesi oksidacije in sinteze ATP. V elektronskem mikroskopu smo v frakcijah plazemskih membran bakterij našli kroglaste delce, podobne tistim v mitohondrijih evkariontskih celic. Tako v bakterijskih celicah, ki so sposobne oksidativne fosforilacije, plazemska membrana igra vlogo, podobno notranji mitohondrijski membrani evkariontskih celic.
Povečanje števila mitohondrijev.
Mitohondrije lahko povečajo njihovo število, zlasti kadar se celice delijo ali ko se poveča funkcionalna obremenitev celice. Mitohondrije se nenehno obnavljajo. Na primer, v jetrih je povprečna življenjska doba mitohondrij približno 10 dni.
Povečanje števila mitohondrijev se pojavi z rastjo in delitvijo sprednjih mitohondrij. To domnevo je prvi izrazil Altman (1893), ki je mitohondrije opisal pod pojmom "bioblasti". Opaziti je mogoče intravitalno delitev, drobljenje dolgih mitohondrij na krajše z zožitvijo, kar spominja na binarni način delitve bakterij.
Resnično povečanje števila mitohondrijev po delitvi je bilo ugotovljeno s preučevanjem vedenja mitohondrijev v živih celicah tkivne kulture. Med celičnim ciklom mitohondriji zrastejo do nekaj mikronov in se nato razdrobijo, razdelijo se na manjša telesa.
Mitohondrije se lahko zlijejo med seboj in se množijo po načelu: mitohondriji iz mitohondrijev.
Avtoprodukcija mitohondrijev.
Organele z dvema membranama celoten sistem avtoreprodukcija. V mitohondrijih in plastidah obstaja DNA, na kateri se sintetizirajo informacije, prenos in ribosomske RNK in ribosomi, ki izvajajo sintezo mitohondrijskih in plastidnih proteinov. Vendar so ti sistemi, čeprav avtonomni, v svojih zmožnostih omejeni.
DNK v mitohondrijih je ciklična molekula brez histonov in tako spominja na bakterijske kromosome. Njihova velikost je približno 7 mikronov, ena ciklična molekula živalskih mitohondrij vključuje 16-19 tisoč nukleotidnih parov DNK. Pri ljudeh mitohondrijska DNK vsebuje 16,5 tisoč bp in je popolnoma dekodirana. Ugotovljeno je bilo, da je mitohondralna DNK različnih predmetov zelo homogena, njihova razlika je le v velikosti intronov in neopisanih regij. Vse mitohondrijske DNK so več kopij, razporejenih v skupinah, grozdih. Torej v enem mitohondriju jeter podgane lahko vsebuje od 1 do 50 cikličnih molekul DNK. Skupna količina mitohondrijske DNK na celico je približno en odstotek. Sinteza DNA mitohondrijev ni povezana s sintezo DNK v jedru.
Tako kot pri bakterijah se mitohondralna DNK zbira v ločenem območju - nukleoidu, njegova velikost je v premeru približno 0,4 mikrona. Dolge mitohondrije lahko vsebujejo od 1 do 10 nukleoidov. Ko je dolg mitohondrij razdeljen, se od njega loči odsek, ki vsebuje nukleoid (podobno kot binarna cepitev bakterij). Količina DNK v posameznih mitohondrijskih nukleoidih lahko niha 10-krat, odvisno od vrste celic.
V nekaterih kulturah v celicah od 6 do 60% mitohondrijev manjka nukleoid, kar je mogoče razložiti s tem, da je delitev teh organelov bolj verjetno povezana z razdrobljenostjo kot s porazdelitvijo nukleoidov.
Kot smo že omenili, se mitohondriji lahko med seboj razdelijo in zlijejo. Ko se mitohondriji združijo med seboj, se lahko zamenjajo njihove notranje komponente.
Pomembno je poudariti, da se rRNA in ribosomi mitohondrijev in citoplazme močno razlikujejo. Če v citoplazmi najdemo ribosome iz 80. let, potem mitohondrijski ribosomi rastlinskih celic spadajo v ribosome 70-ih (sestavljajo jih podenote 30-ih in 50-ih, vsebujejo RNA-je 16-ih in 23-ih, značilne za prokariontske celice), manjši ribosomi (približno 50-ih) pa se nahajajo v mitohondrijah živalskih celic.
Ribosomska RNA mitohondrijev se sintetizira na mitohondrijski DNK. V mitoplazmi se na ribosomih sintetizirajo beljakovine. V nasprotju s sintezo na citoplazemskih ribosomih se ustavi pod delovanjem antibiotika kloramfenikol, ki zavira sintezo beljakovin v bakterijah.
Na mitohondrijskem genomu se sintetizira 22 transportnih RNK. Trojna koda sintetičnega sistema mitohondrijev se razlikuje od tiste, ki se uporablja v hiloplazmi. Kljub prisotnosti na videz vseh komponent, potrebnih za sintezo beljakovin, majhne molekule mitohondrijske DNK ne morejo kodirati vseh proteinov mitohondrijev, le majhen del njih. Torej DNK z velikostjo 15 kb. lahko kodirajo beljakovine s skupno molekulsko maso približno 6x105. Hkrati skupna molekulska teža beljakovin delca celotnega dihalnega ansambla mitohondrije doseže vrednost približno 2x10 6.
Če upoštevamo, da mitohondriji poleg beljakovin oksidativne fosforilacije vključujejo tudi encime cikla trikarboksilne kisline, encime za sintezo DNK in RNK, encime za aktiviranje aminokislin in druge beljakovine, je mogoče razbrati, da lahko za kodiranje teh številnih beljakovin in rRNA in tRNA količino genske informacije v kratka molekula DNA mitohondrijev očitno manjka. Dešifriranje nukleotidnega zaporedja človeške mitohondrijske DNK je pokazalo, da kodira samo 2 ribosomske RNA, 22 prenosnih RNK \u200b\u200bin samo 13 različnih polipeptidnih verig.
Zdaj je dokazano, da je večina mitohondrijskih beljakovin pod genskim nadzorom iz celičnega jedra in se sintetizira zunaj mitohondrij. Večina mitohondrijskih beljakovin se sintetizira na ribosomih v citosolu. Ti proteini imajo posebna signalna zaporedja, ki jih prepoznajo receptorji na zunanji mitohondrijski membrani. Te beljakovine lahko vključimo vanje (glejte analogijo s peroksisomsko membrano) in se nato premaknete na notranjo membrano. Ta prenos se zgodi na stičnih mestih zunanje in notranje membrane, kjer je tak transport opažen. Večina mitohondrijskih lipidov se sintetizira tudi v citoplazmi.
Vse to govori o endosimbiotskem izvoru mitohondrij, da so mitohondriji organizmi, kot so bakterije, ki so v simbiozi z evkariontsko celico.
Kondriomi.
Zbir vseh mitohondrijev v eni celici se imenuje hondriom. Lahko se razlikuje glede na vrsto celic. V mnogih celicah so hondriomi sestavljeni iz razpršenih številnih mitohondrij, enakomerno razporejenih po citoplazmi ali lokaliziranih v skupinah na mestih intenzivnega uživanja ATP. V obeh primerih mitohondrije delujejo ločeno, njihovo skupno delo, po možnosti usklajeno z nekaterimi signali iz citoplazme. Obstaja tudi povsem drugačna vrsta hondrioma, ko namesto majhnih enojnih razpršenih mitohondrijev v celici obstaja en velikanski razvejan mitohondrij.
Takšne mitohondrije najdemo v enoceličnih zelenih algah (npr. Chlorella). Tvorijo kompleksno mitohondrijsko mrežo ali mitohondrijski retikulum (Reticulum miyochondriale). V skladu s kemoosmotsko teorijo je biološki pomen pojava tako velikanske razvejene mitohondrijske strukture, združene v eno po zunanjih in notranjih membranah, da lahko na kateri koli točki na površini notranje membrane tako razvejanega mitohondrija pride do sinteze ATP, ki bo vstopila v katero koli točko citoplazme, kjer v to je potrebno.
V primeru velikanskih razvejanih mitohondrij se lahko na kateri koli točki na notranji membrani kopiči potencial, ki je dovolj za začetek sinteze ATP. S teh položajev je mitohondrijski retikulum kot električni vodnik, kabel, ki povezuje oddaljene točke takega sistema. Mitohondrijski retikulum se je izkazal za zelo uporabnega ne le za majhne gibljive celice, kot je klorela, ampak tudi za večje strukturne enote, kot so miofibrili v skeletnih mišicah.
Znano je, da skeletno mišico sestavlja masa mišičnih vlaken, simplastov, ki vsebujejo veliko jeder. Dolžina takih mišičnih vlaken doseže 40 mikronov, debelina 0,1 mikrona - to je velikanska struktura, ki vsebuje veliko miofibrilov, ki se vse simultano strdijo. Za krčenje se v vsako kontrakcijsko enoto dostavi velika količina ATP-a, na miofibril, ki ga zagotavljajo mitohondriji na ravni z-diskov. Na vzdolžnih ultra tankih odsekih skeletnih mišic v elektronskem mikroskopu so vidni številni majhni zaobljeni odseki mitohondrijev, ki se nahajajo v bližini sarcomerov. Mišični mitohondriji niso majhne kroglice ali palice, ampak so pajki podobne strukture, katerih procesi se vejo in razširijo na dolge razdalje, včasih po celotnem premeru mišičnega vlakna.
V tem primeru razvejenost mitohondrijev obdaja vsako miofibrilo v mišičnem vlaknu in jim oskrbuje ATP, potreben za krčenje mišic. Zato mitohondriji v ravnini z-diska predstavljajo tipičen mitohondrijski retikulum. Takšen sloj ali dno mitohondrijskega retikuluma se ponovi dvakrat za vsak sarcomere, celotno mišično vlakno pa ima na tisoče prečnih "talnih" plasti mitohondrijskega retikuluma. Nitasti mitohondriji se nahajajo med "nadstropji" vzdolž miofibrilov, ki povezujejo te mitohondrijske plasti. Tako je nastala tridimenzionalna slika mitohondrijskega retikuluma, ki prehaja skozi celoten volumen mišičnega vlakna.
Nadalje je bilo ugotovljeno, da med vejami mitohondrijskega retikuluma in nitastih vzdolžnih mitohondrijev obstajajo posebne intermitohondrijske povezave ali stiki (MMC). Nastanejo s tesno prilegajočimi se zunanjimi mitohondrijskimi membranami v stiku z mitohondriji, medmestni membranski prostor in membrane v tem območju imajo povečano gostoto elektronov. Preko teh posebnih tvorb pride do funkcionalne združitve sosednjih mitohondrij in mitohondrijskih retikulumov v en sam, kooperativni energetski sistem. Vse miofibrile v mišičnih vlaknih se sinhronizirajo po celotni dolžini, zato naj bi dobava ATP-ja v katerem koli delu tega kompleksnega stroja potekala tudi sinhrono, in to se lahko zgodi le, če je ogromno število razvejanih mitohondrij-prevodnikov med seboj povezano z uporabo stikov.
Dejstvo, da so intermitohondrijski stiki (MMC) vključeni v energetsko poenotenje mitohondrijev med seboj, je uspelo v kardiomiocitih, srčnih mišičnih celicah.
Kondriomi srčnih mišičnih celic ne tvorijo razvejanih struktur, predstavljajo pa jih številni majhni podolgovati mitohondriji, ki se ne nahajajo v določenem vrstnem redu med miofibrili. Vendar so vsi sosednji mitohondriji povezani med seboj s pomočjo mitohondrijskih stikov iste vrste kot v skeletnih mišicah, le njihovo število je zelo veliko: v povprečju ima en mitohondrij 2-3 MMC, ki povezujejo mitohondrije v eno verigo, kjer je vsaka povezava taka veriga (Streptio mitochondriale) je ločen mitohondrij.
Izkazalo se je, da najdemo intermitohondrijske stike (MMC) kot obvezno strukturo srčnih celic v kardiomiocitih obeh ventriklov in atrijev vseh vretenčarjev: sesalcev, ptic, plazilcev, dvoživk in teleost rib. Poleg tega so MMK našli (vendar v manjšem številu) v srčnih celicah nekaterih žuželk in mehkužcev.
Število MMK v kardiomiocitih se spreminja glede na funkcionalno obremenitev srca. Število MMC narašča s povečanjem telesne aktivnosti živali in obratno, ko obremenitev srčne mišice upade, se število MMC močno zmanjša.