Din aceasta s-a concluzionat că membrana celulelor roșii din sânge este formată din molecule de lipide dispuse în două straturi. Membrane lipidice model „Organizarea moleculară a membranelor biologice”
11 Instituția de învățământ de la bugetul de stat de învățământ profesional superior „Universitatea de Stat de Medicină Saratov numită după. V.I. Razumovsky Ministerul Sănătății al Rusiei”
1. Fiziologie normală: manual / Ed. A.V. Zavyalova, V.M. Smirnova, 2011. – 368 p.
2. Fiziologie normală: manual [N.A. Agadzhanyan, N.A. Barabash, A.F. Belov și colab.] / Ed. prof. V.M. Smirnova. – Ed. a 3-a. – M.: Centrul de Editură „Academia”, 2010. – 480 p.
3. Fiziologia umană / V.F. Kirichuk, O.N. Antipova, N.E. Babichenko, V.M. Golovcenko, E.V. Ponukalina, I.V. Smyshleeva, L.K. Tokaev / Editat de V.F. Kirichuk. – Ed. a II-a. – Saratov: Editura Universității de Medicină din Saratov, 2009. – 343 p.
4. Fiziologia și fiziopatologia sângelui roșu: manual. indemnizatie / N.P. Chesnokova, V.V. Morrison, E.V. Ponukalina, T.A. Nevvazhay; sub general ed. prof. N.P. Cesnokova. – Saratov: Editura Sarat. miere. Universitatea, 2013. – 80 p.
5. Atlas hematologic / S. Lugovskaya, M.E. Poştaş. editia a 3-a. – Moscova – Tver: Editura Triada SRL, 2011. – P. 3–23.
6. Mecanisme celulare și moleculare de reglare a sistemului hemostatic în sănătate și patologie: monografie / B.I. Kuznik. – Chita: Editura Express, 2010. – p. 261–368.
7. Hematologie / Editat de prof. O.A. Rukavitsina, A.D. Pavlova, E.F. Morshchakova și alții - Sankt Petersburg: SRL „D.P.”, 2007. - P. 29–34.
Caracteristicile organizării structurale a membranei eritrocitare
Globulul roșu este înconjurat de o membrană plasmatică, a cărei structură este bine studiată și este identică cu cea a altor celule. Membrana citoplasmatică a globulelor roșii include un strat dublu de fosfolipide, în timp ce proteinele fie „plutesc” pe suprafața membranelor, fie pătrund în lipide, oferind rezistență și vâscozitate membranelor. Aria membranei unui globul roșu este de aproximativ 140 µm2.
Proteinele reprezintă aproximativ 49%, lipidele - 44%, carbohidrații -7%. Carbohidrații sunt legați chimic fie de proteine, fie de lipide și formează glicoproteine și, respectiv, glicolipide.
Cele mai importante componente ale membranei eritrocitare sunt lipidele, inclusiv până la 48% colesterol, 17-28% fosfotidilcolina, 13-25% sfingomielina și o serie de alte fosfolipide.
Fosfotidilcolina membranei eritrocitelor poartă o sarcină neutră și practic nu interacționează cu canalele de Ca2+ încărcate pozitiv, asigurând astfel atrombogenitatea eritrocitelor. Datorită proprietăților precum fluiditatea și plasticitatea, celulele roșii din sânge sunt capabile să treacă prin capilare cu un diametru de ~ 3 μm.
Proteinele membranei celulelor roșii din sânge sunt împărțite în periferice și integrale. Proteinele periferice includ spectrina, anchirina, proteina 4.1, proteina p55, aducina, etc. Grupul de proteine integrale include fracția 3, precum și glicoforinele A, B, C, O, E. Anchirina formează un compus cu p-spectrina. Aproximativ 340 de membrane și 250 de proteine solubile au fost găsite în eritrocite.
Plasticitatea RBC este asociată cu fosforilarea proteinelor membranei, în special a proteinelor din banda 4.1.
Fracția proteică 4.2. - palidina asigură legarea complexului spectrin-actină-anchirina de fracțiunea 3, aparține grupului de proteine transglutaminaze.
Proteinele contractile ale membranei eritrocitare includ p-actina, tropomodulină, stromatina și tropomiozina.
Glicoforinele sunt proteine integrale ale membranei eritrocitare care determină sarcina negativă care favorizează respingerea eritrocitelor unele de altele și din endoteliul vascular.
Proteina 3 este principala proteină actină care reglează defosforilarea eritrocitelor.
După cum am menționat mai sus, membrana eritrocitară este un complex complex, care include lipide, proteine și carbohidrați organizați într-un anumit mod, care formează straturile exterioare, mijlocii și interioare ale membranei eritrocitare.
În ceea ce privește aranjarea spațială a diferitelor componente chimice ale membranei eritrocitare, trebuie remarcat faptul că stratul exterior este format din glicoproteine cu complexe ramificate de oligozaharide, care sunt secțiunile terminale ale antigenelor din sângele grupului. Componentele lipidice ale stratului exterior sunt fosfatidilcolina, sfingomielina și colesterolul neesterificat. Lipidele din stratul exterior al membranei eritrocitare joacă un rol important în asigurarea constanței structurii membranei și a selectivității permeabilității acesteia pentru diferite substraturi și ioni. Împreună cu fosfolipidele, colesterolul reglează activitatea enzimelor legate de membrană prin modificarea vâscozității membranei și este, de asemenea, implicat în modificarea structurii secundare a enzimelor. Raportul molar colesterol/fosfolipide în membranele celulare la oameni și multe mamifere este de 0,9. O modificare ascendentă a acestui raport se observă la bătrânețe, precum și în unele boli asociate cu metabolismul afectat al colesterolului.
O scădere a fluidității membranei eritrocitelor și o modificare a proprietăților sale se observă, de asemenea, cu o creștere a conținutului de sfingomielină,
Stratul dublu al membranei eritrocitare este reprezentat de „cozi” hidrofobe ale lipidelor polare. Stratul dublu lipidic are o fluiditate pronunțată, care este asigurată de un anumit raport între acizii grași saturați și nesaturați ai părții hidrofobe a stratului dublu. Proteinele integrale, care includ enzime, receptori și proteine de transport, sunt active numai dacă sunt situate în partea hidrofobă a stratului dublu, unde dobândesc configurația spațială necesară activității. Prin urmare, orice modificare a compoziției lipidelor membranei eritrocitare este însoțită de o schimbare a fluidității acesteia și de perturbarea funcționării proteinelor integrale.
Stratul interior al membranei eritrocitare, orientat spre citoplasmă, este format din proteinele spectrină și actină. Spectrina este o proteină specifică eritrocitelor; moleculele sale alungite flexibile, care se leagă de microfilamentele de actină și lipidele de pe suprafața interioară a membranei, formează un fel de schelet eritrocitar. Un mic procent de lipide din stratul interior al membranei celulelor roșii din sânge sunt fosfatidiletanolamina și fosfatidilserina. Mobilitatea proteinelor care dețin stratul dublu lipidic depinde de prezența spectrinei.
Una dintre glicoproteinele importante este glicoforina, care este conținută atât pe suprafața exterioară, cât și pe cea interioară a membranelor eritrocitelor. Glicoforina conține o cantitate mare de acid sialic și are o sarcină negativă semnificativă. Este situat inegal in membrana si formeaza zone proeminente din membrana, care sunt purtatoare de determinanti imunologici.
Structura și starea membranei eritrocitelor, vâscozitatea scăzută a hemoglobinei normale oferă proprietăți plastice semnificative eritrocitelor, datorită cărora eritrocitul trece cu ușurință prin capilare, care au jumătate din diametrul celulei în sine și poate prelua o mare varietate de forme. O altă proteină membranară periferică a eritrocitelor este anchirina, care formează un compus cu molecula P-spectrină.
Funcțiile membranei eritrocitare
Membrana eritrocitară asigură reglarea echilibrului electrolitic al celulei datorită transportului de electroliți dependent de energie activă sau difuziei pasive a compușilor de-a lungul gradientului osmotic.
Membrana eritrocitară are canale permeabile la ioni pentru cationii Na+, K+, pentru O2, CO2, Cl- HCO3-.
Transportul electroliților prin membrana eritrocitară și menținerea potențialului de membrană a acesteia este asigurată de sisteme Na+, K+, Ca2+ - ATPaze dependente de energie.
Membrana eritrocitară este foarte permeabilă la apă, cu participarea așa-numitelor căi de proteine și lipide, precum și anioni, compuși gazoși și slab permeabilă la cationii monovalenți de potasiu și sodiu.
Calea proteică a transferului transmembranar de apă este asigurată cu participarea proteinei „banda 3” care se întinde pe membrana eritrocitară, precum și a glicoforinei.
Natura moleculară a căii lipidelor pentru transportul apei prin membrana eritrocitară este practic necunoscută. Trecerea moleculelor de neelectroliți hidrofili mici prin membrana eritrocitară se realizează în același mod ca și transferul apei, datorită căilor proteice și lipide. Transferul de uree și glicerol prin membrana eritrocitară este asigurat de reacții enzimatice.
O trăsătură caracteristică a membranei eritrocitelor este prezența unui sistem activ de transport puternic pentru anioni monovalenți (clor și fluor) și anioni bivalenți (SO42-, PO42-) datorită proteinelor purtătoare.
Transportul de anioni organici prin membrana eritrocitară este asigurat, ca și transportul de anioni anorganici, cu participarea proteinei „banda 3”.
Membrana eritrocitară asigură transportul activ al glucozei, a cărui cinetică este asigurată de dependența Michaelis-Menten. Un rol important în transportul glucozei prin membrana eritrocitară este atribuit polipeptidei din banda 4,5 (proteinele cu un PM de 55 kD sunt posibili produși de descompunere ai polipeptidei din banda 3). S-a sugerat că proteinele care transportă zaharurile în membrana eritrocitară au un mediu lipidic specific.
Distribuția neuniformă a cationilor monovalenți în sistemul eritrocitelor-plasmă sanguină este menținută cu participarea unei pompe Na+ dependentă de energie, care realizează schimbul transmembranar al ionilor de Na+ eritrocitari cu ionii K+ din plasmă sanguină într-un raport de 3:2. Pe lângă schimbul transmembranar Na+/K+ indicat, pompa Na+ realizează cel puțin încă patru procese de transport: schimb Na+ → Na+; K+→K+schimb; intrare monovalentă de ioni de Na+ cuplată cu ieșirea de K+.
Baza moleculară a pompei de Na+ este enzima Na+, K+ -ATPaza - o proteină integrală ferm asociată cu lipidele membranare, constând din 2 subunități polipeptidice cu un PM de 80-100 kDa.
Sistemul de transport are 3 centri care leagă ionii de Na+, localizați pe partea citoplasmatică a membranei. Pe exteriorul membranei de pe sistemul de transport sunt 2 centre de legare pentru ionii K+. Fosfolipidele membranare joacă un rol important în menținerea activității enzimatice ridicate.
Funcționarea pompei de Ca2+ este asigurată de nucleotide, precum și de compuși cu energie înaltă, în principal ATP, CTP, GTP și într-o măsură mai mică GTP și CTP.
Ca și în cazul pompei de Na+, funcționarea pompei de Ca2+ în eritrocite este asociată cu manifestări ale activității Ca2+, Mg2+ -ATPazei. Aproximativ 700 de molecule de Ca2+, Mg2+ -ATPază se găsesc în membrana unui eritrocit.
Împreună cu funcțiile de barieră și de transport, membrana eritrocitară îndeplinește o funcție de receptor.
Prezența receptorilor pentru insulină, endotelină, ceruloplasmină, a2-macroglobulină, receptori α- și β-adrenergici pe membrana eritrocitelor a fost dovedită experimental. Pe suprafața globulelor roșii există receptori pentru fibrinogen, care au o specificitate destul de mare. Celulele roșii din sânge poartă, de asemenea, receptori pentru histamină, TxA2 și prostaciclină pe membrana lor.
Receptorii pentru catecolamine, care reduc mobilitatea, se găsesc în membrana celulelor roșii din sânge acizi grași lipidele membranelor eritrocitelor, precum și stabilitatea osmotică a eritrocitelor.
S-a stabilit o restructurare a structurii membranei eritrocitelor sub influența concentrațiilor scăzute de insulină, hormon de creștere uman și prostaglandine E și E2.
În membranele eritrocitelor, activitatea c-AMP este, de asemenea, ridicată. Odată cu creșterea concentrațiilor de c-AMP în eritrocite (până la 10-6 M), procesele de fosforilare a proteinelor se intensifică, ceea ce duce, la rândul său, la o modificare a gradului de fosforilare și permeabilitate a membranelor eritrocitelor la ioni de Ca2+.
Membrana eritrocitară conține izoantigene diverse sisteme reacții imunologice care determină apartenența la grup a sângelui uman conform acestor sisteme.
Structura antigenică a membranei eritrocitare
Membrana eritrocitară conține diferite antigene de specie, grup și specificitate individuală. Există două tipuri de izoantigene eritrocitare care determină specificitatea de grup a sângelui uman - aglutinogenii A și B. În consecință, în plasmă sau ser se găsesc două tipuri de izoanticorpi - aglutininele α și β. Sângele uman nu conține aceiași aglutinogeni și aglutinine. Întâlnirea și interacțiunea lor poate avea loc în timpul transfuziei de grupe sanguine incompatibile, ducând la dezvoltarea aglutinarii și hemolizei globulelor roșii.
După cum se știe, grupa sanguină I (0) se caracterizează prin absența aglutinogenilor A și B în eritrocite, cu prezența aglutininelor α și β în plasmă sau ser apare la 40-50% dintre oameni din țările central-europene;
Grupa sanguină II (A) se caracterizează prin prezența aglutinogenului A în membrana eritrocitară, în timp ce plasma sanguină conține β aglutinine. Această grupă de sânge este frecventă la 30-40% dintre oameni.
Grupa sanguină III (B) se caracterizează prin prezența aglutinogenului B în membrana eritrocitelor, iar în plasmă sau ser - prin prezența aglutininelor de tip α. Acest grup de sânge apare la aproximativ 10% din populație.
Grupa sanguină IV (AB) se caracterizează prin prezența aglutinogenilor A și B fixați în membrana celulelor roșii din sânge, în timp ce nu există aglutinine naturale α și β în plasma sanguină sau ser. Acest grup de sânge apare la 6% din populație.
Controlul genetic al sistemului antigenic A, B, O al membranelor eritrocitare este reprezentat de genele O, H, A, B, localizate în brațul lung al perechii a 9-a de cromozomi.
Aglutininele α și β aparțin clasei Ig M, sunt anticorpi naturali, se formează la un copil în primul an de viață, ajungând la maximum la 8 - 10 ani.
Al doilea loc între proprietățile antigenice ale membranelor eritrocitare în semnificație clinică este ocupat de sistemul Rh - Hr. Factorul Rh a fost descoperit pentru prima dată în 1940 de K. Landsteiner și A. Wiener se găsește în celulele roșii din sânge la 85% dintre oamenii din rasa albă. La 15% dintre oameni le lipsesc aceste antigene eritrocitare. În prezent, a fost stabilită natura lipoproteică a antigenelor acestui sistem, există aproximativ 20 dintre acestea formând diverse combinații în membrana eritrocitară; Cele mai comune antigene rhesus sunt 6 soiuri: Rh0 (D), rh’ (C), rh’’ (E), Hr0 (d), hr’ (c), hr’’ (e). Cel mai puternic antigen al acestui grup este Rh0 (D).
Anticorpii sistemului Rh și Hr - anti-rhesusaglutininele sunt dobândite, imune, sunt absente în sângele persoanelor Rh (-) din momentul nașterii, sunt sintetizate în timpul primei transfuzii de sânge Rh (+) la un Rh (-). ) primitoare, precum și în timpul primei sarcini a unui fruct Rh (-) femeie (+). În timpul primei sarcini, acești anticorpi sunt sintetizați lent pe parcursul mai multor luni într-un titru mic, fără a provoca complicații grave la mamă și făt. Când o persoană Rh-negativă intră în contact repetat cu celulele roșii Rh-pozitive, este posibil un conflict Rh. Anticorpii sistemului Rh - Hr aparțin clasei Ig G, astfel încât pătrund cu ușurință în bariera placentară, provocând reacții de aglutinare și hemoliză a globulelor roșii fetale, care este însoțită de dezvoltarea icterului hemolitic la nou-născuți. În cazul transfuziei repetate de sânge donator și primitor incompatibil cu antigenele Rh, poate apărea șocul transfuzional.
Link bibliografic
Chesnokova N.P., Ponukalina E.V., Bizenkova M.N. CURTEA 2. CARACTERISTICI ALE STRUCTURII ȘI FUNCȚIILE MEMBRANEI ERITROCITARE // Progrese în știința naturală modernă. – 2015. – Nr. 1-2. – P. 328-331;URL: http://natural-sciences.ru/ru/article/view?id=34842 (data acces: 25/10/2019). Vă aducem în atenție reviste apărute la editura „Academia de Științe ale Naturii”
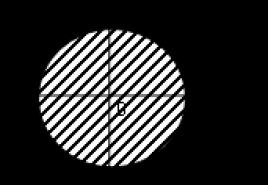
Lipozomii, sau veziculele fosfolipide (veziculele), se obțin de obicei prin umflarea fosfolipidelor uscate în apă sau prin injectarea unei soluții de lipide în apă. În acest caz, are loc auto-asamblarea unei membrane lipidice bimoleculare. Energia Gibbs minimă corespunde formei monolamelare sferice închise a membranei. În acest caz, toate cozile hidrofobe nepolare sunt situate în interiorul membranei și niciuna dintre ele nu intră în contact cu moleculele polare de apă (Fig. 1.11). Cu toate acestea, se obțin mai des lipozomi multilamelari nesferici formați din mai multe straturi bimoleculare - lipozomi multistrat.
Orez. 1.11. Schema structurii unui lipozom cu un singur strat
Straturile bimoleculare individuale ale unui lipozom multistrat sunt separate printr-un mediu apos. Grosimea straturilor lipidice este, în funcție de natura lipidelor, de 6,5 - 7,5 nm, iar distanța dintre ele este de 1,5 - 2 nm. Diametrul lipozomilor multistrat variază de la 60 nm la 400 nm sau mai mult.
Lipozomii cu un singur strat pot fi obținuți prin diferite metode, de exemplu, dintr-o suspensie de lipozomi multistrat prin tratarea lor cu ultrasunete. Diametrul lipozomilor cu un singur strat obținut prin această metodă este de 25-30 nm. Au fost dezvoltate și alte metode pentru producerea de lipozomi cu un singur strat, inclusiv cele cu un diametru de până la 400 nm sau mai mult.
Lipozomii sunt într-un fel un prototip al unei celule. Ele servesc ca model pentru studierea diferitelor proprietăți ale membranelor celulare.
Lipozomii au găsit aplicare directă în medicină. De exemplu, un medicament poate fi închis în lipozomi și utilizat ca microcapsule de fosfolipide pentru a elibera medicamentul către anumite organe și țesuturi. Lipozomii sunt non-toxici (cu selectarea corectă a lipidelor), sunt absorbiți complet de organism și sunt capabili să depășească unele bariere biologice. Astfel, insulina inclusă într-un lipozom este protejată de acțiunea enzimelor digestive. În prezent, se explorează posibilitatea administrării acestui medicament în lipozomi pe cale orală, ceea ce ar putea salva pacienții diabetici de nevoia de injecții sistematice. Se lucrează pentru a dezvolta metode pentru terapia lipozomală pentru tumori, deficit de enzime și ateroscleroză. Se studiază posibilitatea eliberării țintite a unui medicament închis în lipozomi către un organ bolnav sau chiar într-o zonă bolnavă (în special, în zona afectată a inimii).
Pentru a face acest lucru, este atașat de lipozom moleculă proteică-anticorp la antigenul membranar corespunzător al organului țintă. Lipozomii sunt transportați prin fluxul sanguin prin tot corpul și sunt reținuți odată ce sunt în apropierea organului țintă.
În ciuda perspectivelor atractive ale terapiei lipozomale, există încă multe întrebări nerezolvate.
Orez. 1.12. Formarea unei membrane lipidice plane cu două straturi
Membrane lipidice plane cu două straturi (BLM) - alt tip de membrane model. Astfel de membrane sunt produse pe găuri mici cu un diametru de aproximativ 1 mm într-o placă de plastic (de exemplu, fluoroplastic) scufundată într-un mediu apos. O picătură de soluție de lipide (în alcool, cloroform, heptan sau alți solvenți) este aplicată în gaură. Solventul difuzează din soluție în apă, lăsând o peliculă de lipide pe gaură. Acest film se subțiază spontan până când se formează un strat bimolecular de aproximativ 6 nm grosime. Excesul de lipide se adună sub forma unui torus la marginile găurii (Fig. 1.12).
Membranele lipidice plane, împreună cu lipozomii, sunt utilizate pe scară largă ca modele pentru a studia proprietățile electrice ale membranei, permeabilitatea membranei și alte studii științifice. Folosind membrane model, sunt studiate o serie de funcții ale membranelor biologice, inclusiv funcții de barieră (de exemplu, selectivitatea permeabilității - permeabilitate bună pentru apă și permeabilitate slabă pentru ioni). Transportul biologic poate fi modelat prin introducere membrană model molecule purtătoare.
întrebări de testare, sarcini, sarcini
1. Capacitatea electrică specifică a membranei axonilor, măsurată printr-un microelectrod intracelular, s-a dovedit a fi egală cu 0,5 microfarads/cm2. Folosind formula unui condensator plat, estimați grosimea stratului hidrofob al membranei cu o constantă dielectrică de 2.
2. Ce distanță parcurge o moleculă de fosfolipide pe suprafața membranei eritrocitare în 1 secundă ca urmare a difuziei laterale? Coeficientul de difuzie laterală se consideră a fi 10~ 12 m 2 /s. Comparați cu circumferința unei celule roșii din sânge cu un diametru de 8 microni.
3. În timpul tranziției de fază a fosfolipidelor membranare de la starea cristalină lichidă la gel, grosimea stratului dublu se modifică. Cum se va schimba capacitatea electrică a membranei? Cum se vor schimba tensiunile câmp electricîn membrană?
4. Folosind molecule de fosfolipide marcate cu spin, a fost stabilit un gradient de vâscozitate de-a lungul grosimii membranei. Descrie experimentul. Unde este vâscozitatea mai mare: la suprafața membranei sau în centrul acesteia?
teste standard de control al curentului
1.1. Grosimea membranei biologice:
1. 10 A 3,0,1 µm
2. 10 nm 4. 10 um
1.2. Modelul mozaic fluid al unei membrane biologice include:
1. strat proteic, polizaharide și lipide de suprafață!
2. monostratul lipidic și colesterolul
3. bistrat lipidic, proteine, microfilamente
4. dublu strat lipidic
1.3. Partea lipidică a membranei biologice se află în următoarea stare fizică:
1. lichid amorf
2. solid cristalin
3. solid amorf
4. cristal lichid
1.4. Capacitatea electrică specifică a membranei axonului:
1. 0,5 10 -4 F/m 2 3. 0,5 10 -2 F/cm 2
2. 0.5 Yu -2 F/m 2 4. 0.5 10 -12 F/m 2
1.5. Timpul caracteristic de transfer al unei molecule de fosfolipide de la o poziție de echilibru la alta în timpul difuzării lor:
flip-flop lateral
1. 10 -7 – 10 -8 ~1 oră
2. 10 -10 – 10 -12 10 -7 – 10 -8 s
3. 1 – 2 ore 10 – 50 s
1.6. Tranziția de fază a stratului dublu lipidic al membranelor de la starea cristalină lichidă la gel este însoțită de:
1. subtierea membranei
2. grosimea membranei nu se modifică
3. ingrosarea membranei
CAPITOLUL 2. TRANSPORTUL SUBSTANŢELOR PRIN MEMBRANE BIOLOGICE
Sisteme vii la toate nivelurile de organizare - sisteme deschise. Prin urmare, transportul substanțelor prin membranele biologice este conditie necesara viaţă. Procesele metabolice celulare, procesele bioenergetice, formarea de biopotențiale, generarea unui impuls nervos etc. sunt asociate cu transferul de substanțe prin membrane. Încălcarea transportului de substanțe prin biomembrane duce la diferite patologii. Tratamentul implică adesea pătrunderea medicamentelor prin membranele celulare. Eficacitatea unui medicament depinde în mare măsură de permeabilitatea membranei.
Mare valoare a descrie transportul de substante are conceptul de potential electrochimic.
Potențialul chimic al unei substanțe date μ k este o valoare egală numeric cu energia Gibbs pentru un mol din această substanță. Matematic, potențialul chimic este definit ca derivata parțială a energiei Gibbs G în raport cu cantitatea k-ro a substanței, la temperatura constantă T, presiunea P și cantitățile tuturor celorlalte substanțe m 1 (l≠k):
Pentru o soluție diluată cu concentrația de substanță C:
unde μ Q este potențialul chimic standard, egal numeric cu potențialul chimic al unei substanțe date la concentrația sa de 1 mol/l în soluție.
Potențialul electrochimic μ este o valoare egală numeric cu energia Gibbs G pentru un mol dintr-o substanță dată plasată într-un câmp electric.
Pentru soluții diluate
unde F = 96500 C/mol este numărul Faraday, Z este sarcina ionului electrolit (în unități de sarcină elementară), φ este potențialul câmpului electric, T [K] este temperatura.
Transportul substanțelor prin membranele biologice poate fi împărțit în două tipuri principale: pasiv și activ.
Transport activ - transferul de molecule și ioni, care are loc cu cheltuirea energiei chimice în direcția de la valori mai mici la valori mai mari.
În acest caz, moleculele neutre sunt transferate într-o zonă de concentrație mai mare, iar ionii sunt transferați împotriva forțelor care acționează asupra lor din câmpul electric. Astfel, transportul activ realizează transferul de substanțe în direcția opusă transportului, care ar trebui să aibă loc sub influența gradienților (în primul rând concentrația și electrice). Energia este obținută prin hidroliza moleculelor unui compus chimic special - acidul adenozin trifosforic (ATP). Sa stabilit experimental că energia de descompunere a unei molecule de ATP este suficientă pentru a elimina trei ioni de sodiu din exterior și pentru a introduce doi ioni de potasiu în celulă. Diagrama transportului activ este prezentată în Fig. 13.
După ce a captat un ion de potasiu din mediul extern cu un centru activ și un ion de sodiu din mediul intern cu celălalt, sistemul, consumând ATP, se rotește cu 180° în interiorul membranei. Ionul de sodiu ajunge în afara celulei și este separat acolo, iar ionul de potasiu intră și este de asemenea eliberat, după care molecula de proteină își ia poziția inițială și totul începe de la capăt.
Datorită transportului activ, celula menține o concentrație mare de potasiu și o concentrație scăzută de sodiu în ea însăși. În acest caz, ionii se pot deplasa împotriva gradientului lor de concentrație (analogie cu un gaz: pomparea gazului dintr-un vas cu presiune scăzută într-un vas cu presiune mare).
Fig. 13. Schema de transport activ
Transportul activ al substanțelor prin membranele biologice este de mare importanță. Datorită transportului activ, în organism se creează gradienți de concentrație, gradienți de potențial electric, gradienți de presiune etc., care susțin procesele vieții, adică, din punct de vedere al termodinamicii, transportul activ menține corpul într-o stare de neechilibru și susține. viaţă.
Existența transportului activ de substanțe prin membranele biologice a fost dovedită pentru prima dată în experimentele lui Ussing (1949) folosind exemplul transferului ionilor de sodiu prin pielea unei broaște (Fig. 14).
Orez. 14. Schema experimentului lui Ussing (A - ampermetru, V - voltmetru, B - baterie, P - potențiometru)
Camera experimentală a lui Ussing, umplută cu soluție normală de Ringer, a fost împărțită în două părți cu piele de broască proaspăt izolată. În Fig. 14, în stânga este suprafața mucoasă exterioară a pielii, în dreapta este seroasă interioară. S-au observat fluxuri de ioni de sodiu prin pielea unei broaște: de la stânga la dreapta de la suprafața exterioară la suprafața interioară și de la dreapta la stânga - de la suprafața interioară la suprafața exterioară.
O diferență de potențial a apărut pe pielea broaștei care împarte soluția Ringer, partea interioară a pielii având un potențial pozitiv față de cea exterioară. Instalația avea o unitate de compensare a tensiunii, cu ajutorul căreia diferența de potențial de pe pielea broaștei era setată la zero, care era controlată de un voltmetru. În plus, aceeași concentrație de ioni a fost menținută la exterior și în interior. În aceste condiții, dacă transportul ionilor de sodiu prin pielea unei broaște a fost determinat doar prin transport pasiv, atunci fluxurile de ioni de sodiu ar trebui să fie egale între ele și nu ar exista curent în circuit.
Cu toate acestea, s-a descoperit că în condiții experimentale (absența gradienților de potențial electric și de concentrație) curge prin pielea broaștei. curent electric Prin urmare, are loc un transfer într-un singur sens al particulelor încărcate. S-a stabilit că curentul curge prin piele din mediul extern în mediul intern. Folosind metoda atomului marcat, s-a demonstrat că fluxul interior de sodiu este mai mare decât fluxul exterior.
Pentru a face acest lucru, izotopii radioactivi Na 22 au fost incluși în soluția din stânga a camerei experimentale, iar Na 24 în soluția din dreapta. Izotopul Na 22 se descompune odată cu emisia de cuante γ dure. Dezintegrarea Na 24 este însoțită de radiații β moale. Înregistrarea radiațiilor γ - și β - a arătat că fluxul de Na22 este mai mare decât fluxul de Na24. Aceste date experimentale au oferit dovezi de nerefuzat că transportul ionilor de sodiu prin pielea unei broaște nu se supune ecuației de transport pasiv. Prin urmare, are loc transferul activ. Experimentele ulterioare au arătat că epuizarea rezervelor de ATP din pielea broaștei duce la o oprire completă a fluxului unidirecțional al ionilor de sodiu.
3. Scopul activităților elevilor la clasă:
Studentul trebuie sa stie:
1. Rolul membranei în funcționarea celulei.
2. Structura, structura și modelele membranelor.
3. Funcţiile membranei.
4. Proprietăţile fizice ale membranelor.
5. Ecuația lui Fick.
6. Ecuația Nernst-Planck.
7. Tipuri de transport pasiv al particulelor prin membrană.
8. Transportul activ al particulelor prin membrană.
Studentul trebuie să fie capabil să:
1. Explicați structura membranei.
2. Explicați modelele de membrane artificiale.
3. Explicați mecanismul transportului pasiv prin membrană.
4. Explicați mecanismul transportului activ prin membrană.
5. Rezolvați probleme situaționale.
1. Structura membranelor biologice.
2. Modelul mozaic lichid al membranei.
3. Modele cu membrane artificiale.
4. Funcțiile de bază ale membranei celulare.
5. Proprietăţile fizice ale membranelor.
6. Transferul de molecule (atomi) prin membrană. Ecuația lui Fick.
7. Transferul ionilor prin membrane. Ecuația Nernst-Planck.
8. Tipuri de transport pasiv al moleculelor și ionilor prin membrane.
9. Transport activ. Experiența lui Ussing.
10. Rezolvarea problemelor situaționale.
5. Lista de întrebări pentru a verifica nivelul inițial de cunoștințe:
1. Ce sunt membranele biologice?
2. Care este baza membranei?
3. De ce sunt folosite modele de membrane fizico-chimice (artificiale)?
4. Descrieți modelul mozaic lichid al membranei.
5. Ce este difuzia laterală? tranziție flop-flop?
6. Care sunt principalele funcții ale membranei și care sunt acestea?
7. Notați ecuațiile Fick și Nernst-Planck. Ce procese descriu ele?
8. Ce se numește mobilitate?
9. Ce este transportul pasiv? Ce tipuri de transport pasiv există?
10. Ce este transportul activ? Cum se realizează?
11. Care este importanța transportului activ al substanțelor?
12. Explicați fenomenele transferului de materie și de sarcină printr-o membrană.
13. Ce se întâmplă dacă celula este pusă în apă curată?
6 . Lista de întrebări pentru a verifica nivelul final de cunoștințe:
1. Descrieți modelele de membrane lipidice. Unde sunt folosite?
2. Descrieți proprietățile fizice ale membranelor.
3. În timpul tranziției de fază a fosfolipidelor membranare de la starea cristalină lichidă la gel, grosimea stratului dublu se modifică. Cum se va schimba capacitatea electrică a membranei? Cum se va schimba intensitatea câmpului electric din membrană?
4. Aplicați ecuația lui Fick pe o membrană biologică.
5. Scrieți și explicați ecuația Nernst-Planck.
6. Arătați că ecuația Nernst-Planck se reduce la ecuația Fick pentru difuzia particulelor neîncărcate.
7. Descrieți tipurile de transport pasiv.
8. Permeabilitatea membranelor celulare pentru moleculele de apă este de aproximativ 10 ori mai mare decât pentru ioni. Ce se întâmplă dacă concentrația unei substanțe active osmotic (de exemplu, ionii Na+) este crescută într-o soluție apoasă izotonă care conține globule roșii?
9. Descrieți experiența lui Ussing.
7. Rezolvați probleme:
1. Ce distanță parcurge o moleculă de fosfolipide pe suprafața membranei eritrocitare în 1 secundă ca urmare a difuziei laterale? Se ia coeficientul de difuzie laterală egal cu 10 -12 m 2 /s. Comparați cu circumferința unei celule roșii din sânge cu un diametru de 8 microni.
2. Capacitatea electrică specifică a membranei axonilor, măsurată de un microelectrod intracelular, s-a dovedit a fi egală cu 0,5 μF/cm2. Folosind formula pentru un condensator plat, estimați grosimea stratului hidrofob al unei membrane cu o constantă dielectrică de 2.
3. Grosimea stratului dublu la interfața membrană-electrolit este caracterizată de raza Debye δ . Defini δ pentru cazul în care soluția de electrolit care înconjoară membrana conține numai ioni de potasiu cu o concentrație de: 1) 10 -5 mol/l; 2) 10 -2 mol/l.
4. Aflați raza Debye de screening creată de ionii de calciu prezenți în soluție cu o concentrație de 10 -5 mol/l și ionii de sodiu cu o concentrație de 10 -4 mol/l. Cum se va schimba δ, daca solutia contine doar ioni de calciu la o concentratie de 10 -4 mol/l?
5. Raza critică a unui por lipidic dintr-o membrană depinde de tensiunea marginii porului, de tensiunea superficială a membranei și de potențialul membranei. Deduceți o formulă pentru raza critică a porilor. Calculați raza critică a porilor în absența potențialului de membrană. Să presupunem că tensiunea la marginea porului este de 10 -11 N, tensiunea superficială a stratului dublu lipidic este de 0,3 mN/m.
6. Concentrația molară de oxigen în atmosferă cu a= 9 mol/m. Oxigenul difuzează de la suprafața corpului insectelor spre interior prin tuburi numite trahee. Lungimea medie a traheei este de aproximativ h= 2 mm și aria secțiunii sale transversale S= 2∙10 -9 m2. Presupunând că concentrația de oxigen din interiorul insectei ( Cu) este jumătate din concentrația de oxigen din atmosferă, calculați fluxul de difuzie prin trahee. Coeficientul de difuzie a oxigenului D= 10 -5 m 2 /s.
7. Stratul dublu fosfolipidic aseamănă o membrană biologică cu un condensator. Substanța membranei este un dielectric cu o constantă dielectrică ε = 4. Diferența de potențial între suprafețele membranei U= 0,2 V la grosime d= 10 nm. Calculați capacitatea electrică a unei membrane de 1 mm 2 și intensitatea câmpului electric din ea.
8. Aria suprafeței unei celule este aproximativ egală cu S=5∙10 -10 m 2. Capacitatea electrică specifică a membranei (capacitate pe unitate de suprafață) este Curtea= 10 -2 F/m2. În acest caz, potențialul intercelular este egal cu U= 70 mV. Determinaţi: a) cantitatea de sarcină de pe suprafaţa membranei; b) numărul de ioni monovalenți care formează această sarcină.
9. Enzima Na + - K + - ATPaza din membrana plasmatică a eritrocitei a încheiat șase cicluri. Câți ioni de sodiu și potasiu au fost transportați activ? Câtă energie a fost consumată în acest caz dacă hidroliza unui mol de ATP este însoțită de eliberarea a 33,6 kJ? Eficiența procesului de cuplare energetică este considerată 100%.
8. Munca independentă a elevilor:
Folosind manualul de Antonov V.F et al. (§ 15.4.), familiarizați-vă cu metodele fizice de determinare a grosimii membranei.
9. Cronograful sesiunii de antrenament:
1. Moment organizatoric – 5 min.
2. Analiza temei – 50 min.
3. Rezolvarea problemelor situaționale – 40 min.
4. Controlul cunoștințelor curente – 30 min.
5. Rezumatul lecției – 10 min.
10. Lista literaturii educaționale pentru lecție:
1.Remizov A.N., Maksina A.G., Potapenko A.Ya. Fizică medicală și biologică, M., Butarda, 2008, §§ 11.1, 11.2, 11.5, 11.6.
Sânge și celule roșii din sânge. Continuăm să publicăm materiale despre sânge.
Cum arată un globule roșu din sânge? În condiții fiziologice normale în fluxul sanguin, globulele roșii au o formă biconcavă, cu îngroșări uniforme de-a lungul marginilor și o parte centrală mai ușoară - paloarea.
La o examinare optică ușoară, un eritrocit normal colorat în mod obișnuit cu coloranți acizi are forma unui disc cu un diametru de 6,9-7,7 și până la 9,0 microni. În funcție de mărimea lor, celulele roșii din sânge sunt împărțite în micro și macrocite, dar cea mai mare parte a acestora este reprezentată de normocite/discocite.
Proprietățile morfofuncționale ale eritrocitelor
Un eritrocit este o celulă biconcavă anucleată, cu un volum mediu de 90,0 µm 3 și o suprafață de 142 µm 2. Grosimea sa maximă este de 2,4 microni, cea minimă este de 1 microni.
În preparatul uscat, dimensiunea medie a globulelor roșii este de 7,55 microni; 95% din substanța uscată provine din hemoglobina proteică care conține fier și doar 5% din alte substanțe (alte proteine și lipide). Astfel de celule reprezintă majoritatea absolută - peste 85% - a globulelor roșii ale unei persoane sănătoase.
Formele nucleare ale liniei eritrocitelor se disting cu ușurință de cele mai multe celule ale liniei leucocitelor prin absența granulelor în citoplasma lor (erorile sunt posibile numai la identificarea celulelor blastice). Eritroblastele au cromatina nucleară mai granulară și densă.
Cavitatea centrală (paloare) discului eritrocitar reprezintă între 35 și 55% din suprafața acestuia, iar în secțiune transversală eritrocitul are forma unei gogoși, care, pe de o parte, asigură conservarea hemoglobinei și, pe de o parte. alta, permite trecerea eritrocitei chiar si prin cele mai subtiri capilare. Modelele disponibile în prezent ale structurii eritrocitelor corespund ideii proprietăților specifice ale acestei celule, în special învelișul ei, care, în ciuda sensibilității sale la presiunea deformată, rezistă la îndoire și la creșterea suprafeței totale.
Datele din literatură indică faptul că dimensiunea și deformabilitatea membranei eritrocitelor sunt caracteristicile lor cele mai importante, care sunt asociate cu funcționarea normală a acestor celule, inclusiv o capacitate mare de migrare, participarea la procesele metabolice (în primul rând la schimbul de oxigen).
Modificările proprietăților microelastometrice ale eritrocitelor și „transformarea” discocitelor în alte forme morfologice pot fi cauzate de diverși agenți. Astfel, apariția excrescentelor de suprafață duce la o scădere a elasticității membranei, care se poate datora forțelor opuse care apar în chiar procesul de deformare a eritrocitei; deformarea crește odată cu scăderea concentrației de ATP în celule.
Dacă integritatea membranei celulare este încălcată, eritrocitul își pierde forma caracteristică și se transformă într-un sferoplast, care, la rândul său, este hemolizat. Structura membranei eritrocitare (discocitelor) este aceeași în întregime; și în ciuda faptului că depresiuni și umflături pot apărea în diferitele sale părți, modificările presiunii intra- sau extracelulare cu o răspândire de ±15% nu provoacă contracția întregii celule, deoarece are o rezervă semnificativă de „antideformabilitate” . Membrana eritrocitară are suficientă elasticitate pentru a rezista la efectele diferiților factori care apar în timpul circulației eritrocitelor prin fluxul sanguin.
Compoziția membranei eritrocitare include: fosfolipide (36,3%), sfingomieline (29,6%), colesterol (22,2%) și glicolipide (11,9%). Primele două elemente sunt molecule amfifile într-un mediu apos, formând un dublu strat lipidic caracteristic, care este pătruns și de molecule proteice integrale asociate în interiorul eritrocitei cu citoscheletul său.
Lipidele membranare sunt în stare lichidă și au vâscozitate scăzută (de doar 10-100 de ori vâscozitatea apei). Pe suprafața exterioară a membranei există lipide, acid sialic, oligozaharide antigenice și proteine adsorbite; suprafata interioara a membranei este reprezentata de enzime glicolitice, sodiu si calciu, ATPaza, glicoproteine si hemoglobina.
Bistratul lipidic al membranei îndeplinește trei funcții: o funcție de barieră pentru ioni și molecule, o bază structurală pentru funcționarea receptorilor și enzimelor (proteine, glicoproteine, glicolipide) și una mecanică. În implementarea unei funcții respiratorii specializate - transportul de oxigen sau dioxid de carbon - rolul principal este jucat de proteinele membranare, „încorporate” în stratul dublu lipidic. Globulele roșii mature nu sunt capabile să sintetizeze acizi nucleici și hemoglobină; caracteristice acestora nivel scăzut schimb, care asigură o perioadă de viață destul de lungă acestor celule (120 de zile).
Pe măsură ce un globul roșu îmbătrânește, suprafața sa scade, în timp ce conținutul de hemoglobină rămâne neschimbat. S-a stabilit că la vârsta „matură” globulele roșii rămân constante pentru o lungă perioadă de timp compozitia chimica, dar pe măsură ce celulele îmbătrânesc, conținutul de substanțe chimice din ele scade treptat. Citoscheletul eritrocitar este format și controlat de „familii” multigene și de proteine asociate membranei, care organizează domenii membranare specializate care susțin funcția și forma acestei celule înalt specializate.
Potențialul electric al globulelor roșii
Membrana eritrocitară conține 50% proteine, până la 45% lipide și până la 10% carbohidrați. Pe suprafața celulelor intacte, distribuția de „rețea” a sarcinilor este determinată de o glicoproteină care conține acid sialic (neutramic), care determină până la 62% din sarcina negativă de suprafață a celulei.
Se crede că fiecare sarcină electrică corespunde unei molecule din acest acid. Pierderea acidului sialic de la suprafața eritrocitei duce la o scădere a mobilității electroforetice (EPM) a acestuia și la suprimarea transportului de cationi. În consecință, pe suprafața celulelor există un „mozaic” de sarcini determinate de grupuri cationice și anionice, al căror raport determină sarcina electrică totală a eritrocitelor.
Pentru a menține o stare optimă de homeostazie, celulele sanguine trebuie să aibă o încărcare stabilă. Stabilitatea ridicată a EPP este asigurată de un mecanism subtil de reglare a acestuia - echilibrul proceselor de peroxidare a lipidelor (LPO) din membranele eritrocitare și efectul protector al sistemului antioxidant.
S-a stabilit empiric că receptorii pentru anticorpi sunt localizați pe membrana eritrocitelor, iar prezența chiar și a unei mici cantități a acestora la suprafață poate perturba funcțiile fiziologice normale din organism și poate modifica EFP a eritrocitelor. Acest lucru poate afecta nivelul hemoglobinei din acesta din urmă, deoarece conținutul de hemoglobină și EPP sunt strict coordonate.
De asemenea, trebuie avut în vedere că atunci când expunere extremă factori negativi care afectează organismul, produsele de peroxidare a lipidelor afectează proprietățile electrocinetice ale globulelor roșii. La rândul său, acest lucru se reflectă în rata proceselor de peroxid din membranele lor.
Datorită repulsiunii electrostatice („împingere” conform lui Chizhevsky) a celulelor eritrocitare încărcate similar, acestea din urmă se deplasează liber prin vasele de sânge, îndeplinind funcția lor de transport de oxigen. Prin urmare, o încălcare a stabilității sarcinii poate fi considerată un indicator integral al modificărilor patologice din organism.
1. Din aceasta s-a concluzionat că membrana hematiilor este formată din molecule lipidice dispuse în două straturi.
Aparent, această concluzie a lui Gorter și Grendel s-a dovedit a fi corectă numai datorită compensării reciproce a erorilor, dar în termeni istorici această lucrare a fost de mare importanță, deoarece de atunci conceptul unui strat dublu lipidic ca bază structurală a membranelor biologice a devenit dominantă și de fapt s-a dovedit a fi corectă.
Conceptul unei membrane lipidice bimoleculare a fost dezvoltat în continuare de modelul Davson-Danielli sau „sandwich” propus în 1935, în care se presupunea că proteinele acoperă suprafața unui dublu strat lipidic. A fost un model neobișnuit de succes, iar în următorii 30 de ani, numeroase date experimentale, în special cele obținute prin difracție, raze Xși microscopia electronică, a confirmat pe deplin adecvarea acesteia. Cu toate acestea, s-a descoperit apoi că membranele îndeplinesc o mare varietate de funcții, iar pentru a explica acest fenomen, modelul original Davson-Danielli a fost modificat în mod repetat.
Progresul rapid în membranologie, care a dus la formarea conceptelor moderne, a fost realizat în mare parte datorită progreselor în studiul proprietăților proteinelor membranare. Studiile microscopice electronice folosind metoda congelare-clivaj au arătat că particulele globulare au fost încorporate în membrane. Între timp, biochimiștii, folosind detergenți, au reușit să disocieze membranele în starea de „particule” active funcțional. Datele din studiile spectrale au indicat că proteinele membranare au fost caracterizate printr-un conținut ridicat de elice α și că probabil au format globule, mai degrabă decât să fie distribuite ca monostrat pe suprafața stratului dublu lipidic. Proprietățile nepolare ale proteinelor membranare au sugerat prezența unor contacte hidrofobe între proteine și regiunea nepolară internă a stratului dublu lipidic. În același timp, au fost dezvoltate metode care au făcut posibilă dezvăluirea fluidității dublului strat lipidic. Singer și Nicholson au reunit toate aceste idei pentru a crea un model de mozaic fluid. În cadrul acestui model, membrana este reprezentată ca un dublu strat fosfolipidic fluid în care sunt scufundate proteine care difuzează liber. Modelul anterior Davson-Danielli a fost static și a explicat cu succes datele structurale disponibile la acel moment, obținute la o rezoluție destul de scăzută. În același timp, din 1970, s-a acordat multă atenție studiului proprietăților dinamice și relației acestora cu funcțiile membranei. ÎN ultimii ani Modelul mozaic fluid a suferit, de asemenea, modificări, iar acest proces va continua. În special, acum este clar că nu toate proteinele membranei difuzează liber într-un strat dublu lipidic lichid. Există dovezi ale existenței unor domenii laterale în membrana însăși. Rolul citoscheletului este, de asemenea, studiat cu atenție. Devine din ce în ce mai clar că unele regiuni membranare par să difere ca structură de stratul dublu lipidic clasic. Cu toate acestea, în viitorul previzibil, modelul mozaic fluid în diferitele sale modificări va servi drept bază conceptuală pentru multe studii de membrane.
3. Morfologia membranei
Două metode au jucat un rol important în elucidarea morfologiei membranelor: difracția cu raze X și microscopia electronică. Cu ajutorul lor a fost confirmată corectitudinea modelului cu două straturi. Cu toate acestea, trebuie reținut că ambele metode se confruntă cu o serie de limitări atunci când se elucidează o imagine detaliată a organizării moleculare a membranelor.
3.1 Difracția de raze X
Când se studiază probe cristaline foarte ordonate folosind difracția cu raze X, este posibil să se obțină informații structurale de înaltă rezoluție. În cazul medicamentelor prost comandate, capacitățile acestei metode sunt limitate. Unele sisteme membranare specializate au deja o structură regulată și, prin urmare, pot fi studiate prin metode de difracție cu raze X. Un exemplu de acest fel este teaca de mielină a fibrelor nervoase periferice; este o membrană care, înfășurându-se în mod repetat în jurul axonului, formează un sistem regulat de structuri concentrice membranare. Studiile de difracție cu raze X asupra mielinei, efectuate în anii 30, confirmă adecvarea modelului membranei cu două straturi. Aceeași concluzie se trage din studiul segmentului exterior al tijelor retiniene ale vertebratelor, care sunt sisteme membranare ordonate natural, precum și sisteme ordonate artificial care se formează în timpul prăbușirii veziculelor membranare obținute din mitocondrii și eritrocite în condiții de centrifugare. În toate aceste cazuri, a fost observată o distribuție similară a densității electronice în membrană, prezentată în Fig. 1.4.
Pentru a interpreta datele de difracție de raze X, este necesar să se determine nu numai intensitatea reflexiilor, ci și fazele acestora. În cazul sistemelor cu membrane ambalate în mod regulat, sarcina este mult simplificată, deoarece aceste sisteme constau din elemente repetate cu simetrie centrală.

Datele obținute arată că structura tuturor membranelor este similară: au o regiune interioară hidrofobă cu densitate electronică scăzută și două straturi de grupări polare cu densitate mare de electroni. Datele de difracție de raze X obținute pentru diferite membrane diferă doar puțin, în ciuda diferențelor mari în conținutul lor de proteine. Deși datele de difracție cu raze X oferă unele informații despre modul în care cea mai mare parte a proteinelor membranei este localizată în membrană, în general, metoda de difracție cu raze X nu oferă o imagine moleculară detaliată.
Wilkins și colab. au remarcat în 1971 că difracția cu raze X ar putea fi folosită și pentru a studia dispersiile apoase ale membranelor și fosfolipidelor. În acest caz, reflexiile generate de regiunile polare de pe ambele părți ale stratului dublu fac posibilă găsirea grosimii acestuia egală cu distanța dintre capetele polare, iar din reflexiile generate de lanțurile de hidrocarburi ordonate, distanța dintre aceste lanțuri poate fi determinat. Și în acest caz, preparatele membranare obținute din diferite surse au dat un model de difracție similar, ceea ce confirmă universalitatea modelului cu două straturi.
Incapacitatea de a obține o imagine moleculară detaliată folosind metoda difracției limitează utilizarea acestei metode pentru studiul membranelor biologice. Cu toate acestea, poate fi foarte util în studiul sistemelor ordonate lipide-apă.
3.2 Microscopia electronică
Microscopia electronică cu transmisie a secțiunilor subțiri de mielină și, într-adevăr, a tuturor celorlalte membrane, dezvăluie o structură caracteristică cu trei straturi constând din două benzi dense de electroni separate printr-un interval de aproximativ 80 A. Această imagine este obținută în mare parte ca rezultat al tratamentului preparate cu tetroxid de osmiu, utilizate de obicei în această metodă. Robertson a numit structura observată „unitară” pentru a-i sublinia versatilitatea și, deși mecanismele moleculare ale colorării membranelor cu osmiu sunt necunoscute, structura a fost văzută ca susținând validitatea modelului membranei cu două straturi. Este clar, totuși, că atunci când se pregătesc specimene pentru microscopia electronică cu transmisie, membranele pot fi supuse efectelor adverse. În special, se știe că tratamentul cu tetroxid de osmiu duce la pierderi semnificative de proteine din membrana eritrocitară. Și, deși structura cu trei straturi observată în acest caz reflectă într-o oarecare măsură organizarea membranelor bistraturi, informații mai detaliate privind localizarea proteinelor nu pot fi obținute prin această metodă.
Unele informații despre localizarea proteinelor membranare au fost furnizate de noi metode, care au devenit acum „clasice” - metodele de congelare-clivaj și congelare-gravare. În aceste cazuri, preparatele sunt înghețate rapid fără a le expune la vreo influență dăunătoare, ca la obținerea secțiunilor subțiri. Procesul de preparare a medicamentelor include următoarele operații.
După congelare, proba, care este o suspensie de celule sau membrane, este scindată cu un cuțit la temperatură scăzută în vid înalt. Forțele generate în timpul forfecării au ca rezultat formarea unei forfecare care trece prin eșantion. S-a dovedit că atunci când planul tăiat trece prin membrană, aceasta din urmă se împarte în principal de-a lungul regiunii sale de mijloc și se împarte în două jumătăți. Ca rezultat, regiunea internă a membranei este expusă pe planurile de clivaj rezultate.
Dacă este necesar, proba este gravată - se efectuează sublimarea obișnuită a gheții în vid. Acest lucru permite o mai bună vizualizare a structurilor de suprafață ale membranelor celulare.
După aceasta, de pe suprafața expusă se obține o așa-numită replică. Această replică este studiată la microscop electronic. Pentru a obține o replică, platina este mai întâi pulverizată pe probă la un unghi de aproximativ 45° pentru a dezvălui caracteristicile topologice ale medicamentului. Replica din platină primește apoi rezistență mecanică prin acoperirea cu un strat de carbon. După aceasta, medicamentul este dezghețat, replica plutește în sus și este prins folosind o plasă specială.

Cele mai caracteristice structuri observate la studierea membranelor prin metoda îngheț-clivaj sunt numeroase particule intramembranare cu un diametru de 80 până la 100 A, situate în planul clivajelor membranei. De obicei sunt localizați haotic, dar uneori formează grupuri. Numeroase studii au arătat că aceste particule pot fi proteine de membrană. Este curios că microscopia electronică a secțiunilor subțiri nu dezvăluie astfel de structuri. Replicile obținute din cele două jumătăți ale membranei despicate nu sunt întotdeauna complementare topologic. Aceasta înseamnă că unele particule sunt asociate doar cu o jumătate a membranei. Datele de îngheț-clivaj au fost utilizate pe scară largă de Singer și Nicholson pentru a dezvolta modelul mozaic fluid al membranelor, deoarece au arătat în mod convingător că proteinele globulare nu erau localizate doar pe suprafața membranei, ci și în interiorul stratului dublu.
Figura 1.6 prezintă o micrografie electronică a unui preparat de proteolipozomi reconstruiți din fosfatidilcolină de ou și un preparat nefracționat de proteină din banda 3 din membrana eritrocitelor umane; Medicamentul a fost obținut prin congelare - ciobire.
Proteina de bandă 3 este o componentă proteică majoră a membranei celulelor roșii din sânge și este cunoscută că transportă anioni. Dacă veziculele fosfolipide nu conțin această proteină, atunci preparatele de cip congelate rezultate au o suprafață netedă.
Atunci când proteina din banda 3 este încorporată în veziculele fosfolipide, pe suprafețele scindate apar particule intramembranare, practic imposibil de distins de particulele observate în membranele eritrocitelor. Mai mult, la pH 5,5, particulele observate în membrana eritrocitară se agregă, iar această agregare are loc ca urmare a interacțiunii proteinei din banda 3 cu alte două proteine, spectrina și actina.
Acestea din urmă sunt componente ale citoscheletului situat pe suprafața interioară a membranei eritrocitare. Sistemul reconstruit constând din proteina din banda 3 și fosfatidilcolină se comportă în mod similar, cu agregarea particulelor observată în prezența spectrinei și actinei la pH 5,5, dar nu la pH 7,6.

Aceste date au întărit și mai mult ideea proteinelor membranare ca particule globulare care se mișcă liber în planul membranei. În mod interesant, micrografiile statice ale preparatelor obținute prin metoda congelare-clivare au ajutat cercetătorii să studieze proprietățile dinamice ale membranelor. După cum vom vedea, există multe proteine în membrane care nu pot pluti liber în marea lipidelor.
4. Izolarea membranelor
În ultimele trei decenii, a devenit din ce în ce mai clar că marea majoritate a funcțiilor celulare sunt realizate direct de membrane.
Atât celulele vegetale, cât și cele animale sunt împărțite în compartimente, iar multe organele citoplasmatice, așa cum sa arătat în secțiunea 1.1, sunt de natură membranară.
Pe lângă organelele caracteristice majorității celulelor, există și sisteme membranare specializate, cum ar fi reticulul sarcoplasmatic al celulelor musculare, teaca de mielină a fibrelor nervoase periferice, membranele tilacoide ale cloroplastelor și membranele discului din tijele retiniene. Organismele procariote au și membrane, deși nu la fel de dezvoltate ca cele eucariote.
Bacteriile gram-pozitive, cum ar fi Bacillus subtilis, au doar o membrană citoplasmatică, în timp ce bacteriile gram-negative, cum ar fi Escherichia coli, au și o membrană exterioară, situată deasupra unui perete celular subțire de peptidoglican.
Unele organite specializate se găsesc și în celulele procariote. Unii virusuri care sunt patogeni pentru animale, cum ar fi virușii înveliți, au membrane reale, iar astfel de membrane s-au dovedit extrem de interesante de studiat.
Studiul membranelor, de regulă, implică purificarea lor, iar fiecare tip de membrană are propriile condiții pentru izolarea pregătitoare.
Deci, dacă doriți să examinați membrana plasmatică a oricăror celule, atunci trebuie mai întâi să izolați aceste celule din țesut. Apoi, trebuie să găsiți condițiile optime pentru distrugerea celulelor și separarea membranelor de interes de alte componente celulare. Criteriile de puritate ale membranelor izolate merită o atenție deosebită.
4.1 Distrugerea celulelor
Este recomandabil să alegeți o tehnică care vă permite să distrugeți efectiv celulele în sine, menținând în același timp structura membranelor care urmează să fie izolate. Pentru multe celule animale, poate fi utilizată o procedură relativ ușoară, cum ar fi omogenizarea într-un omogenizator Dounce cu pereți de sticlă sau Potter-Elweheim cu un pistil de teflon. În acest caz, celulele sunt distruse din cauza forțelor de forfecare care apar atunci când suspensia este forțată printr-un spațiu îngust dintre pistilul de teflon și peretele de sticlă al omogenizatorului. Cu acest tratament, membrana plasmatică este „smulsă” și conexiunile dintre diferitele organite sunt distruse, menținând în același timp integritatea organelelor în sine. Folosind această procedură, este, de asemenea, posibilă separarea regiunilor specializate ale membranei plasmatice unele de altele, de exemplu, regiunile bazolaterale sau apicale ale membranei celulelor epiteliale. Este de dorit să se lucreze în condiții în care integritatea organelor este menținută pentru a minimiza posibilitatea eliberării enzimelor hidrolitice și pentru a facilita operațiunile ulterioare de separare a membranei.
Pentru a distruge celulele care au un perete, sunt necesare metode mai stricte. Uneori, înainte ca celulele să fie distruse, acestea sunt mai întâi tratate cu enzime care descompun componente ale peretelui celular pentru a facilita distrugerea ulterioară a acestuia. De exemplu, tratamentul cu tampon Tris-EDTA și lizozimă este utilizat pentru a distruge celulele E. coli. Tehnicile mai stricte includ măcinarea celulelor, tratarea lor cu ultrasunete și extrudarea lor. Frecarea se efectuează de obicei în prezența diferitelor materiale abrazive - nisip, oxid de aluminiu sau margele de sticlă. Volumele mici de material pot fi măcinate într-un mojar și pistil, dar pentru volume mai mari ar trebui folosite dispozitive mecanice speciale. Celulele bacteriene adesea distruse cu ajutorul ultrasunetelor. Se crede că în acest caz distrugerea are loc sub influența forțelor tăietoare rezultate din cavitație. Aceleași forțe apar atunci când se presează o suspensie de celule printr-o gaură mică, de exemplu, când se distrug celulele folosind o presă franceză. Există multe varietăți ale metodelor enumerate, iar alegerea lor depinde de caracteristicile sistemului de membrană care este studiat.
Trebuie remarcat faptul că fragmentele de membrană obținute în timpul distrugerii celulelor formează de obicei în mod spontan vezicule. Exemplele includ:
1) microzomi obținuți din membrana plasmatică, reticulul endoplasmatic sau sisteme specializate precum membrana sarcoplasmatică;
2) particule mitocondriale transmise din membrana mitocondrială internă;
3) sinaptozomii formați atunci când terminațiile nervoase sunt rupte în zona contactelor sinaptice;
4) vezicule membranare bacteriene formate din membrana plasmatică a E. coli. Veziculele se formează și din alte sisteme membranare, de exemplu din membranele aparatului Golgi. Dimensiunea lor în majoritatea cazurilor depinde în mare măsură de metoda de distrugere a celulelor. Acest lucru este deosebit de important deoarece dimensiunea veziculelor determină în mare măsură viteza de sedimentare a acestora în timpul centrifugării și comportamentul lor în etapele ulterioare de purificare a membranei. Unele membrane nu formează vezicule, în special membranele suprafețelor laterale ale celulelor animale în contact unele cu altele. Când astfel de celule sunt distruse, o pereche de fragmente de membrană adiacente, ținute împreună de zona de contact, se separă. Prezența unor astfel de contacte împiedică închiderea fragmentelor în vezicule, astfel încât membranele sunt eliberate sub formă de plăci sau structuri de tip panglică.
Alegerea corectă a mediului este, de asemenea, de mare importanță atunci când se distrug celulele. De exemplu, pentru a menține închiderea organelelor membranare, ar trebui să se folosească un mediu care este izosmotic față de conținutul lor intern. Cel mai adesea, se folosește o soluție de zaharoză într-o concentrație de 0,25-0,30 M. În unele cazuri, este mai bine să se folosească sorbitol și manitol. Trebuie remarcat faptul că menținerea izotonicității joacă, de asemenea, un rol important în etapele ulterioare ale izolării pregătitoare a organitelor intacte.
4.2 Separarea membranei
În prezent, centrifugarea este folosită cel mai adesea pentru a separa membranele. Particulele din membrană pot fi clasificate în funcție de viteza lor de sedimentare sau densitatea de plutire. Prima metodă se numește centrifugare zonală și separarea are loc în funcție de valorile S, iar a doua este centrifugarea izopicnică și separarea are loc în condiții de densitate de echilibru. În practică, se utilizează de obicei un hibrid dintre aceste două metode. Figura 1.7 arată poziția unor unități subcelulare pe planul de coordonate „S-g”.
Axa absciselor arată coeficienții de sedimentare ai particulelor, iar axa ordonatelor arată densitatea.

Principiul separării prin viteza de sedimentare poate fi ușor de înțeles prin compararea valorilor S pentru diferite fracții. De exemplu, nucleele au valori S relativ ridicate, i.e. rata lor de sedimentare este semnificativ mai mare decât cea a majorității celorlalte organite subcelulare. Nucleii pot fi peletați selectiv prin centrifugarea omogenatului celular, lăsând toate celelalte organite în supernatant. În același timp, reticulul endoplasmatic neted și aspru nu poate fi separat prin centrifugare zonală.
Diferențele de densitate sunt adesea folosite pentru a izola diferite fracțiuni de membrană dintr-un omogenat celular. În acest scop, centrifugarea se efectuează într-un gradient de densitate. Zaharoza este cel mai adesea folosită pentru a crea un gradient de densitate, dar această metodă are dezavantaje serioase. Pentru a obține densitatea necesară pentru separarea diferitelor fracții de membrană, este necesar să se pregătească soluții cu concentrații mari de zaharoză, care au o vâscozitate mare și sunt, de asemenea, hipertonice. Introducerea organelelor subcelulare într-o soluție hipertonică de zaharoză duce la deshidratarea acestora, iar aducerea ulterioară a soluției în condiții izotonice este adesea însoțită de liză și deteriorarea organelelor. O altă problemă este că multe organele membranare sunt permeabile la zaharoză. Acest lucru poate duce, de asemenea, la distrugerea osmotică a organitelor. Pătrunderea zaharozei în organele membranare separate le poate modifica densitatea efectivă.
Tabelul 1.1. Timpul fizic folosește din ce în ce mai mult alte medii pentru a crea un gradient de densitate. Unele dintre aceste medii sunt enumerate în Tabelul 1.1
Pentru a rezolva aceste probleme, cele mai recente proprietăți ale mediilor de gradient.
1. Ficoll. Un polimer hidrofil al zaharozei, care poate fi utilizat pentru a obține soluții cu o densitate de până la 1,2 g/ml. Principalul său avantaj este presiunea osmotică scăzută a soluțiilor în comparație cu soluțiile cu o concentrație echivalentă de zaharoză , este posibil să se creeze soluții izotonice în intervalul de concentrații datorită includerii suplimentare a zaharozei sau a sărurilor acceptabile fiziologic în mediu. Dezavantajele sunt vâscozitatea ridicată a soluțiilor rezultate și dependența semnificativ neliniară a vâscozității și osmolarității de concentrație. .
2. Metrizamidă. Glucoză triiodo-substituită benzamidă Soluțiile de metrizamidă au o densitate mai mare decât soluțiile Ficoll la aceleași concentrații. Principalul avantaj al soluțiilor de metrizamidă este vâscozitatea lor foarte scăzută, care permite o separare mai rapidă O soluție de metrizamidă de 35% are o osmolaritate aproape fiziologică, astfel încât majoritatea operațiunilor de separare cu membrane pot fi efectuate fără a le expune la soluții hipertonice. Metrizoatul de sodiu este un compus înrudit cu metrizamidă cu proprietăți similare, singura diferență fiind că soluția sa este izotonă la o concentrație de aproximativ 20%. Metrizoatul de sodiu este utilizat în principal pentru izolarea celulelor intacte. Nicodenz este, de asemenea, un derivat al acidului triiodobenzoic, dar are trei catene laterale hidrofile. Când este centrifugat, își formează rapid propriul gradient de densitate; folosit pentru a izola organele subcelulare.
Percoll. O suspensie coloidală de silicagel, ale cărei particule sunt acoperite cu polivinilpirolidonă. Această acoperire reduce efectele toxice ale gelului de silice. Principalul avantaj al Percoll este că nu pătrunde în membranele biologice, iar soluțiile sale au vâscozitate scăzută și osmolaritate scăzută. Datorită dimensiunii mari ale particulelor, centrifugarea soluției Percoll la viteze moderate are ca rezultat formarea unui gradient de densitate. Prin urmare, separarea are loc de obicei foarte repede. Mediul folosit pentru centrifugare poate fi izotonic pe tot volumul său datorită includerii de săruri sau zaharoză. Nu este dificil să se creeze un gradient blând, care să permită o separare foarte eficientă a fracțiilor membranei pe baza densității lor plutitoare.
Sorbitol și manitol. Aceste substanțe sunt uneori folosite în locul zaharozei deoarece, conform datelor publicate, ele pătrund în unele membrane biologice mai puțin bine decât zaharoza.
Rețineți că glicerolul nu este utilizat pentru a crea un gradient de densitate, deoarece nu poate atinge valori suficient de mari de densitate. Sărurile de metale alcaline, cum ar fi CsCl, sunt utilizate numai atunci când sunt necesare soluții de înaltă densitate. Dar trebuie avut în vedere faptul că, în concentrațiile necesare pentru a crea densitatea de echilibru, aceste săruri au adesea un efect dăunător asupra organitelor membranei.
Alte metode sunt, de asemenea, folosite pentru a izola membranele din omogenate celulare, deși nu la fel de frecvent ca centrifugarea.
1. Distribuția fazelor. În acest caz, separarea particulelor de membrană are loc în conformitate cu acestea proprietățile suprafeței. În acest scop, se formează două straturi nemiscibile solutii apoase diferiți polimeri solubili în apă. Un exemplu este un amestec de polietilen glicol dextran și dextran ficoll. Particulele de membrană sunt separate în funcție de afinitatea lor pentru aceste faze. Acestea din urmă pot fi selectate astfel încât să separe membranele în funcție de sarcina lor de suprafață sau de hidrofobicitate.
Electroforeză continuă în flux liber. În acest caz, separarea particulelor are loc în funcție de sarcina lor electrică. Medicamentul de separat este introdus continuu într-un strat subțire de tampon care curge pe un perete vertical. În acest caz, un câmp electric este aplicat perpendicular pe direcția curgerii. Astfel, separarea electroforetică a particulelor are loc în tamponul care curge, care se colectează în partea inferioară a camerei sub formă de fracții separate.
Adsorbție prin afinitate. Separarea se bazează pe o interacțiune biospecifică între componentele membranei și faza solidă. Odată cu descoperirea anticorpilor monoclonali, a devenit posibilă crearea unor tehnici de preparare bazate pe utilizarea componentelor antigenice specifice pentru izolarea membranelor. Anticorpii rezultați pot fi atașați covalent la un suport solid și cu ajutorul lor pot realiza legarea specifică a membranelor corespunzătoare. Această metodă este folosită cel mai adesea pentru izolarea proteinelor membranare. Una dintre problemele care apar aici este legată de selecția condițiilor de eluare a membranei care nu ar provoca denaturarea proteinelor.
Metodă bazată pe utilizarea microgranulelor de silicagel. De obicei, membranele plasmatice nu reprezintă mai mult de 1°7o din masa totală a tuturor membranelor celule eucariote. Prin urmare, izolarea membranelor plasmatice absolut pure este plină de mari dificultăți. O abordare care a fost dezvoltată special pentru izolarea membranelor plasmatice se bazează pe utilizarea microbilor de silicagel cationizate. Aceste granule sunt puternic adsorbite pe suprafața exterioară a membranei plasmatice a celulelor intacte, iar fracțiunea de membrane plasmatice asociată cu granule este ușor separată de-a lungul gradientului de densitate a zaharozei de alte membrane datorită densității mai mari a granulelor. Particularitatea acestei metode este că, în preparatul rezultat, membrana plasmatică cu suprafața sa interioară este transformată în soluție.
4.3 Criterii de puritate pentru fracțiunile de membrană
Poate cel mai obiectiv criteriu pentru puritatea fracției de membrană izolată este prezența în ea a oricărui component unic care este conținut numai în această membrană sau este predominant în ea. De obicei, astfel de componente sunt enzime, care în acest caz sunt numite markeri. O listă a enzimelor marker care sunt utilizate pentru a controla puritatea fracțiilor de membrană este dată în Tabelul 1.2. La determinarea activității enzimei, trebuie luat în considerare faptul că aceasta poate fi într-o formă latentă, de exemplu, datorită faptului că că este localizat pe suprafaţa interioară a veziculelor membranare secretate. Alte probleme asociate cu evaluarea purității membranelor izolate sunt discutate în revizuire. Trebuie remarcat faptul că metodele recomandate în majoritatea cazurilor sunt destul de bine stabilite și standardizate.
În unele cazuri, markerii de membrană mai convenabil nu sunt enzimele, ci receptorii specifici pentru lectine, hormoni, toxine sau anticorpi. Dacă sistemele studiate sunt bine caracterizate, atunci puritatea fracției de membrană poate fi judecată după compoziția sa proteică, determinată folosind electroforeza pe gel de poliacrilamidă în prezența dodecil sulfatului de sodiu. De exemplu, membrana exterioară a bacteriilor gram-negative are un set caracteristic de polipeptide care nu se găsesc în membrana citoplasmatică.
Tabelul 1.2 Markeri utilizați pentru a controla puritatea fracțiilor membranare izolate din celulele de mamifere "
| Fracția de membrană | Enzima marker |
| Membrane plasmatice | 5 "-Nucleotidaza |
| Fosfodiesteraza alcalina | |
| Na */K + -ATPaza (bazolateral- |
|
| membrana epitelială | |
| celule) | |
| Adenilat ciclază (bazal | |
| membrana hepatocitară) | |
| Aminopeptidaza (membrană | |
| epiteliul marginii perie) | |
| Mitocondriile (interne | Citocrom c oxidaza |
| membrană) | succinat-citocrom c-oxido- |
| reductaza | |
| Mitocondriile (exterioare | Monoaminoxidaza |
| membrană) | |
| Lizozomi | Fosfataza acidă |
| 0-galactoză | |
| Peroxizomii | Catalaza |
| Urat oxidaza | |
| D-aminoacid oxidaza | |
| Membrane ale dispozitivului | Galactosiltransferaza |
| Golgi | |
| Endoplasmatic | Glucozo-6-fosfataza |
| reticul | Colina fosfotransferaza |
| NADPH-citocrom c-oxido- | |
| reductaza | |
| Citosol | Lactat dehidrogenază |
Alte criterii după care poate fi apreciată puritatea membranelor includ morfologia lor, evidențiată de microscopia electronică și caracteristicile compoziției lor chimice. De exemplu, fracțiile reprezentând membrana plasmatică, aparatul Golgi sau mitocondriile pot fi identificate prin morfologia lor. În unele cazuri, medicamentul se caracterizează prin conținutul său de colesterol. De exemplu, membranele mitocondriale conțin mult mai puțin colesterol decât membranele aparatului Golgi și membranele plasmatice.
Există molecule de detergent pe micelă. O gamă destul de limitată de detergenți este utilizată în studiile membranelor. În tabel 1 le prezintă pe cele care sunt utilizate cel mai des pentru solubilizarea și reconstrucția membranelor. Acești detergenți se caracterizează prin valori CMC destul de mari (10-4-10-2 M) și prin faptul că aparțin categoriei așa-numiților detergenți moi, adică astfel de...

Formarea bistratului este proprietate specială molecule lipidice și se realizează chiar și în afara celulei. Cele mai importante proprietăți ale unui strat dublu: - capacitatea de auto-asamblare - fluiditate - asimetrie. 1.2. Deși proprietățile de bază ale membranelor biologice sunt determinate de proprietățile stratului dublu lipidic, majoritatea funcțiilor specifice sunt asigurate de proteinele membranei. Majoritatea dintre ele pătrund în stratul dublu sub forma unui singur...