Come vengono chiamate le strutture interne dei mitocondri. La struttura e le funzioni dei mitocondri e dei plastidi
mitocondri
Nelle cellule dei tessuti animali, i mitocondri furono trovati nel 1882 e nelle piante solo nel 1904 (nelle antere delle ninfee). È stato possibile stabilire funzioni biologiche dopo separazione e purificazione della frazione mediante centrifugazione frazionata. Contengono il 70% di proteine \u200b\u200be circa il 30% di lipidi, una piccola quantità di RNA e DNA, vitamine A, B 6, B 12, K, E, acidi folici e pantotenici, riboflavina, vari enzimi. I mitocondri hanno una doppia membrana, l'esterno isola l'organello dal citoplasma e le forme interne emergono dal crista. L'intero spazio tra le membrane è riempito con una matrice (Fig. 13).
La funzione principale dei mitocondri è la partecipazione alla respirazione cellulare. Il ruolo dei mitocondri nella respirazione fu istituito nel 1950-1951. Un complesso sistema enzimatico del ciclo di Krebs è concentrato sulle membrane esterne. Durante l'ossidazione dei substrati respiratori, viene rilasciata energia che viene immediatamente accumulata nelle molecole formate dell'ADP, principalmente ATP, durante la fosforilazione ossidativa che si verifica nelle criste. L'energia immagazzinata nei composti macroergici viene utilizzata in futuro per soddisfare tutte le esigenze della cellula.
La formazione di mitocondri nella cellula si verifica continuamente dai microrganismi, più spesso la loro presenza è associata alla differenziazione delle strutture di membrana della cellula. Possono essere ripristinati nella cella dividendoli e germogliandoli. I mitocondri non sono durevoli, la loro aspettativa di vita è di 5-10 giorni.
I mitocondri sono stazioni "energetiche". Concentrano l'energia, che viene immagazzinata negli "accumulatori" di energia - molecole di ATP e non dissipata nella cellula. La violazione della struttura dei mitocondri porta all'interruzione del processo respiratorio e, di conseguenza, alla patologia del corpo.
Apparato del Golgi.Apparato del Golgi (un sinonimo - dictiosomi) è una pila di 3-12 appiattiti, chiusi, circondati da un doppio disco a membrana chiamato serbatoi, dai bordi dei quali numerose bolle sono sganciate (300-500). La larghezza dei serbatoi è di 6-90 A, lo spessore delle membrane è di 60-70 A.
L'apparato del Golgi è un centro per la sintesi, l'accumulo e l'isolamento dei polisaccaridi, in particolare della cellulosa, ed è coinvolto nella distribuzione e nel trasporto intracellulare delle proteine, nonché nella formazione di vacuoli e lisosomi. Nella cellula vegetale, è stato possibile tracciare la partecipazione dell'apparato del Golgi nell'aspetto della placca mediana e la crescita della membrana cellula-cellulosa delle cellule.
L'apparato del Golgi è maggiormente sviluppato durante il periodo di vita cellulare attiva. Con il suo invecchiamento, si atrofizza gradualmente e poi scompare.
Lisosomi.lisosomi - corpi arrotondati piuttosto piccoli (circa 0,5 micron di diametro). Sono rivestiti con una membrana lipoide-proteica. I contenuti dei lisosomi sono numerosi enzimi idrolitici che svolgono la funzione di digestione intracellulare (lisi) di proteine \u200b\u200bmacromolecole, acidi nucleici, polisaccaridi. La loro funzione principale è la digestione delle singole sezioni del protoplasto cellulare (autofagia - auto-divorante). Questo processo procede a causa di fagocitosi o pinocitosi. Il ruolo biologico di questo processo è duplice. In primo luogo, è protettivo, perché con una carenza temporanea di prodotti di riserva, la cellula supporta la vita a causa di proteine \u200b\u200bcostituzionali e altre sostanze, e in secondo luogo, c'è un rilascio da organelli in eccesso o usurati (plastidi, mitocondri, ecc.) La membrana del lisosoma impedisce il rilascio di enzimi nel citoplasma altrimenti, sarebbe completamente digerito da questi enzimi.
In una cellula morta, i lisosomi vengono distrutti, gli enzimi si trovano nella cellula e tutto il suo contenuto viene digerito. Non resta che il guscio di pecto-cellulosa.
I lisosomi sono prodotti dell'attività dell'apparato del Golgi, vescicole staccate da esso, in cui questo organoide ha accumulato enzimi digestivi.
Sferosomy - corpi arrotondati proteina-lipoidi 0,3-0,4 micron. Con ogni probabilità, sono derivati \u200b\u200bdell'apparato del Golgi o del reticolo endoplasmatico. Nella loro forma e dimensione ricordano i lisosomi. Poiché gli sferosomi contengono fosfatasi acida, sono probabilmente correlati ai lisosomi. Alcuni autori ritengono che sferosomi e lisosomi siano equivalenti tra loro, ma molto probabilmente solo per origine e forma. C'è un'ipotesi sulla loro partecipazione alla sintesi dei grassi (A. Frey-Wissling).
ribosomi - organoidi molto piccoli, il loro diametro è di circa 250 A. In forma sono quasi sferici. Alcuni di essi sono attaccati alle membrane esterne del reticolo endoplasmatico, altri sono in uno stato libero nel citoplasma. Una cellula può contenere fino a 5 milioni di ribosomi. I ribosomi si trovano nei cloroplasti e nei mitocondri, dove sintetizzano parte delle proteine \u200b\u200bdi cui sono costruiti questi organoidi e gli enzimi che funzionano in essi.
La funzione principale è la sintesi di proteine \u200b\u200bspecifiche in base alle informazioni provenienti dal nucleo. La loro composizione: proteine \u200b\u200be acido ribonucleico ribosomiale (RNA) in proporzioni uguali. La loro struttura è di piccole e grandi subunità formate da ribonucleotide.
Microtubuli.microtubuli - derivati \u200b\u200bpeculiari del reticolo endoplasmatico. Trovato in molte cellule. Il loro stesso nome parla della loro forma - uno o due, situati in parallelo, tubuli con una cavità all'interno. Diametro esterno entro 250A. Le pareti dei microtubuli sono costruite con molecole proteiche. I filamenti del fuso sono formati da microtubuli durante la divisione cellulare.
Il nucleo
Il nucleo fu scoperto in una cellula vegetale da R. Brown nel 1831. Si trova al centro della cellula o vicino alla membrana cellulare, ma è circondato dal citoplasma su tutti i lati. Nella maggior parte dei casi, c'è un nucleo nella cellula, diversi nuclei si trovano nelle cellule di alcune alghe, oltre ai funghi. Le alghe verdi non cellulari hanno centinaia di nuclei. Cellule multinucleate di batteri non articolati. Non ci sono nuclei nelle cellule di batteri e alghe blu-verdi.
La forma del nucleo è molto spesso vicina alla forma di una palla o di un'ellisse. Dipende dalla forma, dall'età e dalla funzione della cellula. Nella cellula meristematica, il nucleo è grande, di forma rotonda e occupa 3/4 del volume cellulare. Nelle cellule parenchimali dell'epidermide con un grande vacuolo centrale, il nucleo ha una forma lenticolare e viene spinto insieme al citoplasma alla periferia della cellula. Questo è un segno di una cellula specializzata, ma già invecchiata. Una cellula priva di un nucleo può vivere solo per un breve periodo. Le cellule libere dal nucleare dei setacci sono cellule viventi, ma non vivono a lungo. In tutti gli altri casi, le cellule libere dal nucleare sono morte.
Il nucleo ha un doppio guscio, attraverso i pori in cui il contenuto
i nuclei (nucleoplasma) possono comunicare con il contenuto del citoplasma. Le membrane di membrana del nucleo sono dotate di ribosomi e comunicano con le membrane del reticolo endoplasmatico della cellula. Uno o due nucleoli e cromosomi si trovano nel nucleoplasma. Il nucleoplasma è un sistema colloidale di Zola, che ricorda una gelatina addensata in consistenza. Secondo biochimici russi (I. Zbarsky, et al.), Il nucleo contiene quattro frazioni di proteine: proteine \u200b\u200bsemplici - globuline 20%, desossiribonucleoproteine \u200b\u200b- 70%, proteine \u200b\u200bacide - 6% e proteine \u200b\u200bresidue 4%. Sono localizzati nelle seguenti strutture nucleari: proteine \u200b\u200bdel DNA (proteine \u200b\u200balcaline) nei cromosomi, proteine \u200b\u200bdell'RNA (proteine \u200b\u200bacide) nei nucleoli, in parte nei cromosomi (durante la sintesi dell'RNA messaggero) e nella membrana nucleare. Le globuli formano la base del nucleoplasma. Le proteine \u200b\u200bresidue (natura non specificata) formano una membrana nucleare.
La maggior parte delle proteine \u200b\u200bdi base sono proteine \u200b\u200balcaline complesse di desossiribonucleoproteine, che sono basate sul DNA.
Molecola di DNAMolecola di DNA - polinucleotide ed è costituito da nucleotidi. Il nucleotide contiene tre componenti: una molecola di zucchero (desossiribosio), una molecola di azoto e una molecola di acido fosforico. Il desossiribosio è collegato alla base azotata glicosidica e all'acido fosforico - un legame etereo. Nel DNA, in varie combinazioni ci sono solo 4 varietà di nucleotidi che differiscono tra loro nelle basi azotate. Due di loro (adenina e guanina) appartengono a composti azotati purinici e citosina e timina a pirimidina. Le molecole di DNA non si trovano sullo stesso piano, ma sono costituite da due filamenti a spirale, ad es. due catene parallele intrecciate l'una attorno all'altra formano una molecola di DNA. Sono legati tra loro tramite un legame idrogeno tra basi azotate e le basi puriniche di una catena attaccano le basi pirimidiniche dell'altra (Fig. 14). La struttura e la chimica della molecola del DNA sono state scoperte da scienziati inglesi (Crick) e americani (Watson) e pubblicate nel 1953. Questo punto è considerato come l'inizio dello sviluppo della genetica molecolare. Il peso molecolare del DNA è di 4-8 milioni, il numero di nucleotidi (varie opzioni) è fino a 100 mila. La molecola di DNA è molto stabile, la sua stabilità è garantita dal fatto che per tutta la sua lunghezza ha lo stesso spessore - 20A (8A - la larghezza della base di pirimidina + 12A - la larghezza della base di purina). Se il fosforo radioattivo viene introdotto nel corpo, l'etichetta verrà rilevata in tutti i composti contenenti fosforo, ad eccezione del DNA (Levy, Sickewitz).
Le molecole di DNA sono portatrici dell'eredità, perché la loro struttura codifica le informazioni sulla sintesi di proteine \u200b\u200bspecifiche che determinano le proprietà del corpo. Possono verificarsi cambiamenti sotto l'influenza di fattori mutageni (radiazioni radioattive, potenti, agenti chimici, alcaloidi, alcoli, ecc.).
Molecola di RNAMolecole di acido ribonucleico (RNA) significativamente meno molecole di DNA. Queste sono singole catene di nucleotidi. Esistono tre tipi di RNA: ribosomiale, il più lungo, che forma numerosi anelli, informazioni (matrice) e trasporto, il più breve. L'RNA ribosomiale è localizzato nei ribosomi del reticolo endoplasmatico e rappresenta l'85% dell'RNA totale della cellula.
L'RNA informativo nella sua struttura ricorda una foglia di trifoglio. La sua quantità è del 5% dell'RNA totale nella cellula. È sintetizzato nei nucleoli. Il suo assemblaggio avviene nei cromosomi durante il periodo interfase. La sua funzione principale è il trasferimento di informazioni dal DNA ai ribosomi, dove avviene la sintesi proteica.
L'RNA di trasporto, come è ora stabilito, è un'intera famiglia di composti correlati nella struttura e nella funzione biologica. Ogni cellula vivente, secondo una stima approssimativa, contiene 40-50 singoli RNA di trasporto e il loro numero totale in natura, date le differenze di specie, è enorme. (Acad. V. Engelhardt). Sono chiamati trasporto perché le loro molecole sono impegnate nel mantenimento del trasporto del processo intracellulare di sintesi proteica. In combinazione con aminoacidi liberi, li trasportano ai ribosomi nella catena proteica in costruzione. Queste sono le molecole di RNA più piccole, composte in media da 80 nucleotidi. Localizzato nella matrice del citoplasma e costituisce circa il 10% dell'RNA cellulare
L'RNA contiene quattro basi azotate, ma a differenza del DNA, l'uracile è al posto della timina anziché della timina.
La struttura dei cromosomi.I cromosomi furono scoperti per la prima volta alla fine del XIX secolo dai classici della citologia Fleming e Strasburger (1882, 1884) e dal ricercatore di cellule russo I.D. Chistyakov li ha scoperti nel 1874.
Il principale elemento strutturale dei cromosomi è il nucleo. Hanno una forma diversa. Questi sono bastoncini dritti o curvi, corpi ovali, palle, le cui dimensioni variano.
A seconda della posizione del centromero, si distinguono cromosomi diretti, uguali e disuguali. La struttura interna dei cromosomi è mostrata in Fig. 15, 16. Va notato che la desossiribonucleoproteina è un monomero del cromosoma.
Nel cromosoma delle desossiribonucleoproteine, il 90-92%, di cui il 45% è DNA e il 55% è proteina (istone). In una piccola quantità nel cromosoma, anche l'RNA è rappresentato (informativo).
Il cromosoma ha anche una pronunciata struttura trasversale - la presenza di aree ispessite - dischi, risalenti al 1909. erano chiamati geni. Questo termine è stato proposto dallo scienziato danese Johansen. Nel 1911, lo scienziato americano Morgan ha dimostrato che i geni sono le principali unità ereditarie e sono distribuiti nei cromosomi in un ordine lineare e, quindi, il cromosoma ha sezioni qualitativamente diverse. Nel 1934, lo scienziato americano Pinter ha dimostrato la discontinuità della struttura morfologica dei cromosomi e la presenza di dischi nei cromosomi, e i dischi sono luoghi in cui si accumula il DNA. Questo è servito come l'inizio della creazione di mappe cromosomiche, che indicavano la posizione (locus) della posizione del gene che determina l'uno o l'altro tratto dell'organismo. Un gene è un sito a doppia elica del DNA che contiene informazioni sulla struttura di una singola proteina. Questa è una sezione di una molecola di DNA che determina la sintesi di una singola molecola proteica. Il DNA non partecipa direttamente alla sintesi proteica. Contiene e memorizza solo informazioni sulla struttura della proteina.
La struttura del DNA, composta da diverse migliaia di 4 nucleotidi consecutivi, è un codice di ereditarietà.
Il codice dell'ereditarietà. Sintesi proteica.Il primo messaggio sul codice del DNA è stato scritto dal biochimico americano Nirenberg nel 1961 a Mosca al Congresso biochimico internazionale. L'essenza del codice DNA è la seguente. Ogni amminoacido corrisponde a una porzione della catena del DNA di tre nucleotidi adiacenti (triplette). Quindi, ad esempio, un sito costituito da TT-T (una tripletta di 3 nucleotidi contenenti timina) corrisponde all'amminoacido lisina, una tripletta A (adenina) - C (citosina) - A (adenina) - cisteina, ecc. Supponiamo che un gene sia rappresentato da una catena di nucleotidi disposti nel seguente ordine: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G. Avendo suddiviso questa serie in triplette, decifreremo immediatamente quali aminoacidi e in quale ordine saranno localizzati nella proteina sintetizzata.
Il numero di possibili combinazioni di 4 nucleotidi disponibili di tre è 4 × 64. Sulla base di questi rapporti, il numero di diverse triplette in eccesso sarà sufficiente per fornire informazioni sulla sintesi di numerose proteine \u200b\u200bche determinano sia la struttura che le funzioni del corpo. Per la sintesi proteica, una copia esatta di queste informazioni viene inviata ai ribosomi sotto forma di RNA messaggero. Oltre all'i-RNA, un gran numero di molecole di vari acidi ribonucleici di trasporto (t-RNA), ribosomi e un numero di enzimi sono coinvolti nella decodifica e nella sintesi. Ognuno dei 20 aminoacidi si lega al T-RNA, una molecola con una molecola. Ognuno dei 20 aminoacidi ha il suo t-RNA. I T-RNA hanno gruppi chimici in grado di "riconoscere" il loro aminoacido, scegliendolo tra gli aminoacidi disponibili. Questo accade con l'aiuto di enzimi speciali. Avendo riconosciuto il suo amminoacido, il t-RNA entra in un composto con esso. Un ribosoma è attaccato all'inizio della catena (molecola) dell'i-RNA, che, muovendosi lungo l'i-RNA, si collega tra loro nella catena polipeptidica esattamente quegli aminoacidi il cui ordine è codificato dalla sequenza nucleotidica di questo I-RNA. Di conseguenza, si forma una molecola proteica, la cui composizione è codificata in uno dei geni.
nucleoli- una parte strutturale integrale del nucleo. Questi sono corpi sferici. Sono molto mutevoli, cambiano forma e struttura, appaiono e scompaiono. Ce ne sono uno, due. Per ogni pianta un certo numero. I nuclei scompaiono quando la cellula si prepara alla divisione e poi riappaiono; sono apparentemente coinvolti nella sintesi di acidi ribonucleici. Se il nucleolo viene distrutto da un raggio focalizzato di raggi X o raggi ultravioletti, la divisione cellulare viene soppressa.
Il ruolo del nucleo nella vita cellulare. Il nucleo funge da centro di controllo della cellula: dirige l'attività cellulare e contiene portatori di ereditarietà (geni) che determinano le caratteristiche di un dato organismo. Il ruolo del nucleo può essere identificato se, usando tecniche microchirurgiche, lo rimuove dalla cellula e ne osserva le conseguenze. Una serie di esperimenti che hanno dimostrato un ruolo importante nella regolazione della crescita cellulare è stata effettuata da Hemmerling sull'alga verde unicellulare Acetobularia. Questa alga raggiunge un'altezza di 5 cm, sembra un fungo, ha la somiglianza di "radici" e "zampe". Sopra termina con un grande "cappello" a forma di disco. La cellula di questa alga ha un nucleo situato nella parte basale della cellula.
Hammerling ha scoperto che se la gamba viene tagliata, la parte inferiore continua a vivere e il cappuccio viene completamente rigenerato dopo l'operazione. La parte superiore, priva del nucleo, sopravvive per un po 'di tempo, ma alla fine muore, non potendo ripristinare la parte inferiore. Di conseguenza, il nucleo dell'acetobularia è necessario per le reazioni metaboliche alla base della crescita.
Il nucleo promuove la formazione della parete cellulare. Ciò può essere illustrato da esperimenti con l'alga Voucheria e Spyrogyra. Rilasciando il contenuto di cellule dai filamenti tagliati in acqua, possiamo ottenere grumi di citoplasma con uno, con diversi nuclei e senza nuclei. Nei primi due casi, la membrana cellulare si è formata normalmente. In assenza di un nucleo, la shell non si è formata.
Negli esperimenti di I.I.Gerasimov (1890) con spirogyra, è stato scoperto che le cellule con un doppio nucleo raddoppiano la lunghezza e lo spessore del cloroplasto. Nelle cellule prive di nucleare, il processo di fotosintesi continua, si forma l'amido di assimilazione, ma il processo della sua idrolisi viene attenuato, il che è spiegato dall'assenza di enzimi idrolitici che possono essere sintetizzati nei ribosomi solo in base al DNA del nucleo. La vita di un protoplasto senza nucleo è inferiore e di breve durata. Negli esperimenti di I.I. Le cellule di spirogyra prive di nucleare di Gerasimov sono vissute 42 giorni e sono morte. Una delle funzioni più importanti del nucleo è quella di fornire al citoplasma l'acido ribonucleico, necessario per la sintesi delle proteine \u200b\u200bnella cellula. La rimozione del nucleo dalla cellula porta a una graduale riduzione del contenuto di RNA nel citoplasma e un rallentamento della sintesi proteica in esso.
Il ruolo più importante del nucleo è nel trasferimento di tratti da cellula a cellula, da organismo a organismo, e lo fa nel processo di fissione nucleare e della cellula nel suo insieme.
Divisione cellulare.Le cellule si moltiplicano per divisione. In questo caso, due cellule figlie con lo stesso set di materiale ereditario racchiuso nei cromosomi della cellula madre sono formate da una cellula. Nelle cellule somatiche, i cromosomi sono rappresentati da due, i cosiddetti cromosomi omologhi, in cui sono posati geni allelici (portatori di caratteri opposti, ad esempio, colore bianco e rosso dei petali di piselli, ecc.), Segni di due coppie parentali. A questo proposito, nelle cellule somatiche del corpo vegetale, esiste sempre una doppia serie di cromosi, indicata con 2p. I cromosomi hanno una personalità pronunciata. La quantità e la qualità dei cromosomi è una caratteristica di ogni specie. Quindi, nelle cellule di fragole l'insieme diploide di cromosi è 14, (2n), l'albero di mele è 34, il topinambur è 102, ecc.
Mitosi (cariocinesi) - la divisione cellulare somatica è stata descritta per la prima volta da E. Russov (1872) e I.D. Chistyakov (1874). La sua essenza sta nel fatto che due cellule figlie con lo stesso insieme di cromosi si formano dalla cellula madre per divisione: il ciclo cellulare è composto da interfase e mitosi propriamente detta. Il metodo di microautoradiografia ha scoperto che l'interfase più lunga e più difficile è il periodo del nucleo "a riposo", perché durante questo periodo si raddoppia il materiale nucleare. L'interfase è divisa in tre fasi:
Q1 - presintetico (la sua durata è di 4-6 ore);
S - sintetico (10-20 ore);
Q2 - postsintetico (2-5 ore).
Durante la fase Q1, sono in corso i preparativi per la riduplicazione del DNA. E nella fase S, il DNA è ridotto, la cellula raddoppia la fornitura di DNA. Nella fase Q2 si formano gli enzimi e le strutture necessarie per innescare la mitosi. Pertanto, nell'interfase, le molecole di DNA nei cromosomi sono suddivise in due filoni identici e l'RNA delle informazioni è assemblato sulla loro matrice. Quest'ultimo contiene informazioni sulla struttura di proteine \u200b\u200bspecifiche nel citoplasma e nel nucleo, ciascuno dei filamenti di DNA completa la metà mancante della sua molecola. In questo processo di raddoppio (riduzione), si manifesta una caratteristica unica del DNA, che consiste nella capacità del DNA di riprodursi accuratamente. Le molecole di DNA figlia risultanti vengono automaticamente ottenute da copie esatte della molecola madre, perché durante la riduplicazione, le basi complementari (АТ; Г-Ц; ecc.) dall'ambiente si uniscono ogni metà.
Nella fase della divisione mitotica, i cromosomi raddoppiati diventano evidenti. In metafase, si trovano tutti nella zona equatoriale, situata in una riga. Si formano i filetti del mandrino (dai microtubuli che si collegano tra loro). Il guscio del nucleo e del nucleolo scompaiono. I cromosomi ispessiti sono suddivisi longitudinalmente in due cromosomi figlie. Questa è l'essenza della mitosi. Fornisce una distribuzione accurata delle molecole di DNA raddoppiate tra le cellule figlie. Ciò garantisce la trasmissione di informazioni ereditarie crittografate nel DNA.
In anafase, i cromosomi figlie iniziano a spostarsi su poli opposti. Al centro compaiono i primi frammenti della membrana cellulare (fragmoblast).
Nella telofase, i nuclei si formano nelle cellule figlie. Il contenuto della cellula madre (organello) è distribuito tra le cellule figlie risultanti. La membrana cellulare è completamente formata. Questo termina la citochinesi (Fig. 17).
Meiosi - Divisione di riduzione fu scoperto e descritto negli anni '90 del secolo scorso da V.I. Belyaev. L'essenza della divisione è che da una cellula somatica contenente un set di cromosomi 2n (doppio, diploide), si formano quattro cellule aploidi, con "n", una mezza serie di cromosi. Questo tipo di divisione è complessa e si compone di due fasi. Il primo è la riduzione dei cromosomi. I cromosomi doppi si trovano nella zona equatoriale in coppia (due cromosomi omologhi paralleli). In questo momento, può verificarsi coniugazione (adesione) per cromosi, incrocio (incrocio) e, di conseguenza, si verifica uno scambio di siti di cromosi. Di conseguenza, parte dei geni dei cromosomi paterni diventa parte delle cromosi materne e viceversa. L'aspetto di entrambi i cromosomi a seguito di ciò non cambia, ma la loro composizione qualitativa diventa diversa. Le eredità paterne e materne vengono ridistribuite e mescolate.
Nell'anafase della meiosi, i cromosomi omologhi con l'aiuto dei filamenti del fuso divergono ai poli, in cui, dopo un breve periodo di riposo (i filamenti scompaiono, ma il setto tra i nuovi nuclei non si forma), inizia il processo di mitosi - una metafase in cui tutti i cromosomi si trovano su un piano e si verifica la loro divisione longitudinale su cromosomi figlie. Con l'anafase della mitosi con l'aiuto di un fuso, divergono ai poli, dove si formano quattro nuclei e, di conseguenza, quattro cellule aploidi. Nelle cellule di alcuni tessuti durante il loro sviluppo sotto l'influenza di determinati fattori, si verifica una mitosi incompleta e il numero di cromosi nei nuclei raddoppia a causa del fatto che non divergono ai poli. Come risultato di tali violazioni di natura naturale o artificiale, sorgono tetraploidi e poliploidi. Con l'aiuto della meiosi, si formano le cellule germinali: gameti, nonché spore, elementi di riproduzione sessuale e asessuata delle piante (Fig. 18).
L'amitosi è una fissione diretta del nucleo. Con l'amitosi, non si forma un fuso di divisione e il guscio del nucleo non si decompone, come nella mitosi. In precedenza, l'amitosi era considerata una forma primitiva di divisione. È stato ora stabilito che è associato al degrado del corpo. È una versione semplificata di una fissione più complessa del kernel. L'amitosi si verifica nelle cellule e nei tessuti del nucello, dell'endosperma, del parenchima dei tuberi, dei piccioli fogliari, ecc.
mitocondrio - È un organello a spirale, rotondo, allungato o ramificato.
Il concetto di mitocondri fu proposto per la prima volta da Benda nel 1897. I mitocondri possono essere rilevati nelle cellule viventi usando il contrasto di fase e la microscopia di interferenza sotto forma di granuli, granuli o filamenti. Queste sono strutture abbastanza mobili che possono muoversi, fondersi l'una con l'altra, condividere. Se colorati con metodi speciali in cellule morte sotto microscopia ottica, i mitocondri sembrano piccoli grani (granuli), distribuiti diffusamente nel citoplasma o concentrati in alcune zone specifiche.
Come risultato della distruzione di glucosio e grassi in presenza di ossigeno, nei mitocondri viene generata energia e la materia organica viene convertita in acqua e anidride carbonica. In questo modo, gli organismi animali ricevono l'energia di base necessaria per la vita. L'energia si accumula nell'adenosina trifosfato (ATP) e, più precisamente, nei suoi legami macroergici. La funzione dei mitocondri è strettamente correlata all'ossidazione dei composti organici e all'uso dell'energia rilasciata durante il loro decadimento per la sintesi delle molecole di ATP. Pertanto, i mitocondri sono spesso chiamati stazioni energetiche cellulari o organelli della respirazione cellulare. L'ATP funge da fornitore di energia, trasferendo uno dei suoi gruppi terminali di fosfati ricchi di energia in un'altra molecola e si trasforma in ADP.
Si ritiene che in evoluzione i mitocondri fossero microrganismi procariotici che sono diventati simbiotici nel corpo di una cellula antica. Successivamente, divennero vitalmente necessari, il che fu associato ad un aumento del contenuto di ossigeno nell'atmosfera terrestre. Da un lato, i mitocondri hanno rimosso l'ossigeno tossico in eccesso per la cellula e, dall'altro, hanno fornito energia.
Senza i mitocondri, la cellula non è praticamente in grado di utilizzare l'ossigeno come sostanza che fornisce energia e può soddisfare il proprio fabbisogno energetico solo attraverso processi anaerobici. Pertanto, l'ossigeno è veleno, ma il veleno è vitale per la cellula e l'eccesso di ossigeno è altrettanto dannoso della sua carenza.
I mitocondri possono cambiare la loro forma e spostarsi in quelle aree della cellula in cui il loro bisogno è più elevato. Quindi, nei cardiomiociti, i mitocondri si trovano vicino alle miofibrille, nelle cellule dei tubuli renali vicino alle sporgenze basali, ecc. La cellula contiene fino a mille mitocondri e il loro numero dipende dall'attività della cellula.
I mitocondri hanno una dimensione trasversale media di 0,5 ... 3 micron. A seconda delle dimensioni, si distinguono i mitocondri piccoli, medi, grandi e giganti (formano una rete ramificata - il reticolo mitocondriale). Le dimensioni e il numero di mitocondri sono strettamente correlati all'attività della cellula e al suo consumo di energia. Sono estremamente variabili e, a seconda dell'attività cellulare, del contenuto di ossigeno, delle influenze ormonali, possono gonfiarsi, cambiare il numero e la struttura delle criste, variare in numero, forma e dimensioni, nonché l'attività enzimatica.
La densità apparente dei mitocondri, il grado di sviluppo della loro superficie interna e altri indicatori dipendono dal fabbisogno energetico della cellula. Nei linfociti ci sono solo pochi mitocondri e nelle cellule del fegato ce ne sono 2 ... 3 mila.
I mitocondri sono costituiti da una matrice, una membrana interna, uno spazio perimitocondriale e una membrana esterna. La membrana mitocondriale esterna separa l'organello dall'ialoplasma. Di solito ha contorni lisci ed è chiuso in modo che sia una sacca a membrana.
La membrana esterna dall'interno è separata da uno spazio perimitocondriale con una larghezza di circa 10 ... 20 nm. La membrana mitocondriale interna limita l'effettivo contenuto interno dei mitocondri - la matrice. La membrana interna forma numerose sporgenze verso l'interno dei mitocondri, che hanno l'aspetto di creste piatte o cristi.
Le criste possono assumere la forma di placche (trabecolari) e tubuli (multivicolari al taglio) di forma e sono dirette longitudinalmente o trasversalmente rispetto ai mitocondri.
Ogni mitocondrio è riempito con una matrice, che nelle microfotografie elettroniche sembra più densa del citoplasma circostante. La matrice mitocondriale è omogenea (omogenea), a volte a grana fine, di varie densità di elettroni. Si rivelano sottili filamenti con uno spessore di circa 2 ... 3 nm e granuli con una dimensione di circa 15 ... 20 nm. I filamenti di matrice sono molecole di DNA e piccoli granuli sono ribosomi mitocondriali. La matrice contiene enzimi, un DNA a singolo filamento, ciclico, ribosomi mitocondriali, molti ioni Ca 2+.
Il sistema autonomo di sintesi proteica dei mitocondri è rappresentato da molecole di DNA esenti da istoni. Il DNA è corto, ha la forma di un anello (ciclico) e contiene 37 geni. A differenza del DNA nucleare, non vi sono praticamente sequenze di nucleotidi non codificanti. Le caratteristiche della struttura e dell'organizzazione riuniscono il DNA mitocondriale con il DNA delle cellule batteriche. Sul DNA mitocondriale vengono sintetizzati vari tipi di molecole di RNA: informativo, trasferimento (trasporto) e ribosomiale. Informazioni L'RNA dei mitocondri non è soggetto a giunzioni (tagliare sezioni che non portano un carico di informazioni). Le piccole dimensioni delle molecole di DNA mitocondriale non sono in grado di determinare la sintesi di tutte le proteine \u200b\u200bmitocondriali. La maggior parte delle proteine \u200b\u200bmitocondriali sono sotto il controllo genetico del nucleo cellulare e sono sintetizzate nel citoplasma, poiché il DNA mitocondriale è debolmente espresso e può fornire la formazione di solo una parte degli enzimi ossidativi della catena della fosforilazione. Il DNA mitocondriale codifica non più di dieci proteine \u200b\u200blocalizzate nelle membrane e sono proteine \u200b\u200bstrutturali responsabili della corretta integrazione dei singoli complessi proteici funzionali delle membrane mitocondriali. Anche le proteine \u200b\u200bche svolgono funzioni di trasporto sono sintetizzate. Un tale sistema di sintesi proteica non fornisce tutte le funzioni dei mitocondri, quindi l'autonomia dei mitocondri è limitata e relativa.
Nei mammiferi, i mitocondri durante la fecondazione vengono trasmessi solo attraverso l'uovo e lo sperma introduce il DNA del nucleo nel nuovo organismo.
Nella matrice dei mitocondri si formano ribosomi che differiscono dai ribosomi citoplasmatici. Partecipano alla sintesi di un numero di proteine \u200b\u200bmitocondriali non codificate dal nucleo. I ribosomi mitocondriali hanno un numero di sedimentazione di 60 (in contrasto con il citoplasmatico con un numero di sedimentazione di 80). Il numero di sedimentazione è il tasso di deposizione durante la centrifugazione e l'ultracentrifugazione. Per struttura, i ribosomi mitocondriali sono vicini ai ribosomi degli organismi procariotici, ma sono più piccoli e sensibili a determinati antibiotici (cloramfenicolo, tetraciclina, ecc.).
La membrana mitocondriale interna ha un alto grado di selettività nel trasporto di sostanze. Enzimi della catena ossidativa della fosforilazione, proteine \u200b\u200btrasportatrici di elettroni, ATP, ADP, piruvato e altri sistemi di trasporto vicini l'uno all'altro sono attaccati alla sua superficie interna. Come risultato della stretta disposizione degli enzimi sulla membrana interna, è assicurata un'alta coniugazione (interconnessione) dei processi biochimici, che aumenta la velocità e l'efficienza dei processi catalitici.
La microscopia elettronica rivela particelle di funghi che sporgono nel lume della matrice. Hanno un'attività ATP-sintetica (forma ATP dall'ADP). Il trasporto di elettroni avviene lungo la catena respiratoria localizzata nella membrana interna, che contiene quattro grandi complessi enzimatici (citocromi). Quando gli elettroni passano attraverso la catena respiratoria, gli ioni idrogeno vengono pompati dalla matrice nello spazio perimitocondriale, il che assicura la formazione di un gradiente protonico (pompa). L'energia di questo gradiente (differenze nella concentrazione di sostanze e nella formazione del potenziale di membrana) viene utilizzata per la sintesi di ATP e il trasporto di metaboliti e ioni inorganici. Le proteine \u200b\u200bportatrici contenute nella membrana interna trasportano attraverso di essa fosfati organici, ATP, ADP, aminoacidi, acidi grassi, acidi tri e bicarbossilici.
La membrana mitocondriale esterna è più permeabile alle sostanze a basso peso molecolare, poiché contiene numerosi canali proteici idrofili. I complessi recettori specifici si trovano sulla membrana esterna, attraverso la quale le proteine \u200b\u200bdella matrice vengono trasportate nello spazio perimitocondriale.
In termini di composizione chimica e proprietà, la membrana esterna è vicina ad altre membrane intracellulari e al plasmolemma. Contiene enzimi che metabolizzano i grassi, attivano (catalizzano) la conversione di ammine, ammino ossidasi. Se gli enzimi della membrana esterna rimangono attivi, questo è un indicatore della sicurezza funzionale dei mitocondri.
Esistono due sottocompartimenti autonomi nei mitocondri. Lo spazio perimitocondriale, o la camera mitocondriale esterna (sottocompartimento esterno), si forma a causa della penetrazione dei complessi proteici dell'ialoplasma, mentre il sottocompartimento interno (matrice mitocondriale) è parzialmente formato a causa dell'attività sintetica del DNA mitocondriale. Il sottocompartimento interno (matrice) contiene DNA, RNA e ribosomi. Ha un alto livello di ioni Ca 2+ rispetto allo ialoplasma. Gli ioni idrogeno si accumulano nel sottocompartimento esterno. L'attività enzimatica dei sottocompartimenti esterni ed interni, la composizione delle proteine \u200b\u200bsono molto diverse. Il compartimento interno ha una densità di elettroni superiore a quella esterna.
Marcatori mitocondriali specifici sono gli enzimi citocromo ossidasi e succinato deidrogenasi, la cui rilevazione consente la caratterizzazione quantitativa dei processi energetici nei mitocondri.
La funzione principale dei mitocondri - Sintesi ATP. Inizialmente, nello ialoplasma, gli zuccheri (glucosio) vengono distrutti in acidi lattici e piruvici (piruvato) con la sintesi simultanea di una piccola quantità di ATP. Come risultato della glicolisi di una molecola di glucosio, vengono utilizzate due molecole di ATP e ne vengono formate quattro. Pertanto, solo due molecole di ATP costituiscono il bilancio positivo. Questi processi si verificano senza ossigeno (glicolisi anaerobica).
Tutte le fasi successive della produzione di energia avvengono nel processo di ossidazione aerobica, che fornisce la sintesi di una grande quantità di ATP. In questo caso, la materia organica viene distrutta in CO 2 e acqua. L'ossidazione è accompagnata dal trasferimento dei protoni ai loro accettatori. Queste reazioni vengono eseguite utilizzando un numero di enzimi del ciclo dell'acido tricarbossilico, che si trovano nella matrice mitocondriale.
I sistemi di trasferimento di elettroni e la relativa fosforilazione dell'ADP (fosforilazione ossidativa) sono integrati nelle membrane cristali. In questo caso, ha luogo il trasferimento di elettroni da una proteina accettratrice ad un'altra e, infine, il loro legame con l'ossigeno, a seguito del quale si forma l'acqua. Allo stesso tempo, parte dell'energia rilasciata durante tale ossidazione nella catena di trasferimento degli elettroni viene immagazzinata come legame macroergico durante la fosforilazione dell'ADP, che porta alla formazione di un gran numero di molecole di ATP, il principale equivalente di energia intracellulare. Il processo di fosforilazione ossidativa si verifica sulle membrane delle malattie mitocondriali utilizzando le proteine \u200b\u200bdella catena di ossidazione e l'enzima di fosforilazione della sintetasi ADP ATP che si trova qui. Come risultato della fosforilazione ossidativa, si formano 36 molecole di ATP da una molecola di glucosio.
Ad alcuni ormoni e sostanze sui mitocondri ci sono recettori specializzati (di affinità). La triiodotironina normalmente accelera l'attività sintetica dei mitocondri. L'interleuchina-1 e le alte concentrazioni di triiodotironina disconnettono le catene ossidative della fosforilazione, causando gonfiore mitocondriale, che è accompagnato da un aumento della formazione di energia termica.
I nuovi mitocondri sono formati da divisione, costrizione o erba. In quest'ultimo caso si formano protomitocondri, che aumentano gradualmente di dimensioni.
I protomitocondri sono piccoli organelli con membrane esterne ed interne. La membrana interna non ha o contiene criteri sottosviluppati. L'organella è caratterizzata da bassi livelli di fosforilazione aerobica. Con la formazione di una costrizione, il contenuto dei mitocondri viene distribuito tra due nuovi organelli abbastanza grandi. Con qualsiasi metodo di riproduzione, ciascuno dei mitocondri di nuova formazione ha il suo genoma.
I mitocondri più vecchi vengono distrutti dall'autolisi (auto-digestione da parte della cellula utilizzando i lisosomi) per formare autolisosomi. Dall'autolisosoma si forma un corpo residuo. Con una digestione completa, il contenuto del corpo residuo, costituito da sostanze organiche a basso peso molecolare, viene escreto dall'esocitosi. Con digestione incompleta, i resti di mitocondri possono accumularsi nella cellula sotto forma di corpi stratificati o granuli con nipofuscina. I sali di calcio insolubili si accumulano nella parte dei mitocondri con la formazione di cristalli - calcificazioni. L'accumulo di prodotti di degenerazione mitocondriale può portare alla distrofia cellulare.
La struttura e la funzione dei mitocondri è una questione piuttosto complessa. La presenza di organelli è caratteristica di quasi tutti gli organismi nucleari, sia per gli autotrofi (piante capaci di fotosintesi) sia per gli eterotrofi, che sono quasi tutti animali, alcune piante e funghi.
Lo scopo principale dei mitocondri è l'ossidazione delle sostanze organiche e il successivo uso dell'energia rilasciata a seguito di questo processo. Per questo motivo, gli organelli hanno anche un secondo nome (non ufficiale): stazioni di energia cellulare. A volte vengono chiamati "plastidi del catabolismo".
Cosa sono i mitocondri
Il termine è di origine greca. Tradotta, questa parola significa "thread" (mitos), "seed" (condrion). I mitocondri sono organoidi permanenti che sono di grande importanza per il normale funzionamento delle cellule e rendono possibile l'esistenza dell'intero organismo.

Le "stazioni" hanno una struttura interna specifica, che varia a seconda dello stato funzionale dei mitocondri. La loro forma può essere di due tipi: ovale o oblunga. Quest'ultimo ha spesso un aspetto ramificato. Il numero di organelli in una cellula varia da 150 a 1500.
Un caso speciale sono le cellule germinali. Nello sperma è presente solo un organello a spirale, mentre i gameti femminili contengono centinaia di migliaia di mitocondri in più. Nella cellula, gli organelli non sono fissati in un punto, ma possono muoversi lungo il citoplasma, combinandosi tra loro. La loro dimensione è di 0,5 micron, la lunghezza può raggiungere i 60 micron, mentre la cifra minima è di 7 micron.

Determinare le dimensioni di una "centrale elettrica" \u200b\u200bnon è un compito facile. Il fatto è che se visti al microscopio elettronico, solo una parte dell'organello entra nella fetta. Accade così che i mitocondri a spirale abbiano diverse sezioni che possono essere prese per strutture separate e indipendenti.
Solo un'immagine tridimensionale consentirà di scoprire l'esatta struttura cellulare e capire se stiamo parlando di 2-5 organoidi separati o di uno con una forma complessa di mitocondri.
Caratteristiche strutturali
La membrana mitocondriale è composta da due strati: esterno e interno. Quest'ultimo include varie escrescenze e pieghe che hanno una forma a foglia e tubolare.
Ogni membrana ha una composizione chimica speciale, una certa quantità di determinati enzimi e uno scopo specifico. Il guscio esterno dall'interno è separato da uno spazio intermembrana con uno spessore di 10-20 nm.
La struttura dell'organello nella figura con le didascalie sembra molto chiara.

La struttura dei mitocondri
Guardando il diagramma della struttura, è possibile fare la seguente descrizione. Lo spazio viscoso all'interno dei mitocondri è chiamato matrice. La sua composizione crea un ambiente favorevole per il flusso dei processi chimici necessari al suo interno. Contiene granuli microscopici che promuovono reazioni e processi biochimici (ad esempio, accumulano ioni glicogeno e altre sostanze).
La matrice contiene DNA, coenzimi, ribosomi, t-RNA, ioni inorganici. ATP sintasi e citocromi si trovano sulla superficie dello strato interno del guscio. Gli enzimi contribuiscono a processi come il ciclo di Krebs (CTK), la fosforilazione ossidativa, ecc.
Pertanto, il compito principale dell'organoide viene svolto sia dalla matrice che dal lato interno del guscio.
Funzione mitocondriale
Lo scopo delle "centrali elettriche" può essere caratterizzato da due compiti principali:
- produzione di energia: in essi vengono eseguiti processi di ossidazione con il successivo rilascio di molecole di ATP;
- archiviazione di informazioni genetiche;
- partecipazione alla sintesi di ormoni, aminoacidi e altre strutture.
Il processo di ossidazione e produzione di energia si svolge in più fasi:


Disegno schematico della sintesi di ATP
Vale la pena notare: a seguito del ciclo di Krebs (ciclo dell'acido citrico), le molecole di ATP non si formano, le molecole vengono ossidate e l'anidride carbonica viene rilasciata. Questo è un passaggio intermedio tra glicolisi e catena di trasporto degli elettroni.
Tabella "Funzioni e struttura dei mitocondri"

Cosa determina il numero di mitocondri in una cellula
Il numero prevalente di organoidi si accumula vicino a quelle parti della cellula dove c'è bisogno di risorse energetiche. In particolare, un gran numero di organelli viene raccolto nell'area delle miofibrille, che fanno parte delle cellule muscolari che ne assicurano la contrazione.

Nelle cellule germinali maschili, le strutture sono localizzate attorno all'asse del flagello - si presume che la necessità di ATP sia dovuta al costante movimento della coda del gamete. La disposizione dei mitocondri nei protozoi, che usano speciali ciglia per il movimento, sembra che gli organelli si accumulino sotto la membrana alla base esattamente nello stesso modo.
Per quanto riguarda le cellule nervose, la localizzazione dei mitocondri viene osservata vicino alle sinapsi attraverso le quali vengono trasmessi i segnali del sistema nervoso. Nelle cellule che sintetizzano le proteine, gli organelli si accumulano nelle zone ergastoplasma - forniscono energia che fornisce questo processo.
Chi ha scoperto i mitocondri
La struttura cellulare prese il nome nel 1897-1898 grazie a C. Brand. Otto Wagburg è stato in grado di dimostrare la relazione della respirazione cellulare con i mitocondri nel 1920.
conclusione
I mitocondri sono il componente più importante di una cellula vivente, fungendo da stazione di energia che produce molecole di ATP, garantendo così i processi di attività cellulare.
Il lavoro dei mitocondri si basa sull'ossidazione dei composti organici, con conseguente generazione di potenziale energetico.
Membrana esterna
Membrana interna
matrice m-on, matrice, crista. ha contorni lisci, non forma sporgenze o pieghe. Rappresenta circa il 7% dell'area di tutte le membrane cellulari. Il suo spessore è di circa 7 nm, non è collegato ad altre membrane del citoplasma ed è chiuso a se stesso, quindi è un sacchetto di membrana. La membrana esterna è separata dall'interno spazio intermembrana circa 10-20 nm di larghezza. La membrana interna (circa 7 nm di spessore) limita l'effettivo contenuto interno di mitocondri,
la sua matrice o mitoplasma. Una caratteristica della membrana interna dei mitocondri è la loro capacità di formare numerose sporgenze all'interno dei mitocondri. Tali sporgenze il più delle volte hanno la forma di creste piatte o criste. La distanza tra le membrane nella crista è di circa 10-20 nm. Spesso le criste possono ramificare o formare processi a forma di dito, piegarsi e non avere un orientamento pronunciato. Nei protozoi, alghe unicellulari, in alcune cellule delle piante e degli animali superiori, le escrescenze della membrana interna sembrano tubi (croste tubulari).
La matrice mitocondriale ha una struttura omogenea a grana fine, a volte filamenti sottili raccolti in una palla (circa 2-3 nm) e in essa vengono rilevati granuli di circa 15-20 nm. Ora è noto che i filamenti della matrice mitocondriale sono molecole di DNA nella composizione del nucleoide mitocondriale e che i piccoli granuli sono ribosomi mitocondriali.
Funzione mitocondriale
1. Nei mitocondri avviene la sintesi di ATP (vedi. Fosforilazione ossidativa)
Intermembrana pH ~ 4, matrice pH ~ 8 | il contenuto proteico in m: il 67% è la matrice, il 21% è la scoperta della miniera, il 6% è l'interno della miniera e il 6% è nella miniera
Handrioma - un singolo sistema mitocondriale
m-na esterno: i pori porinici consentono il passaggio fino a 5 kD | m-na interno: m-well impermeabile alla cardiolipina per ioni |
produzione internazionale: gruppi enzimatici nucleotidi fosforilati e zuccheri nucleotidici
m interno-on:
matrice: enzimi metabolici - ossidazione dei lipidi, ossidazione dei carboidrati, ciclo dell'acido tricarbossilico, ciclo di Krebs
Origine dai batteri: l'ameba Pelomyxa palustris non contiene unità dagli eucarioti, vive in simbiosi con i batteri aerobici | proprio DNA | Processi simili a OX
DNA mitocondriale
Divisione dei mioclondri
replicato
in interfase | la replica non è associata alla fase S | durante il ciclo cl, i mitociti sono una volta divisi in due, formando una costrizione, la costrizione prima all'interno m-not | ~ 16,5 kb | anello codifica 2 rRNA 22 tRNA e 13 proteine \u200b\u200b|
trasporto di proteine: segnale peptide | ricciolo anfifilico | recettore del riconoscimento mitocondriale |
Fosforilazione ossidativa
Circuito di trasferimento di elettroni
ATP sintasi
nelle cellule del fegato, vivono circa 20 giorni di divisione mitocondriale per formazione di costrizione
16569 bp \u003d 13 proteine, 22tRNA, 2pRNA | liscia esterna m-na (porine - permeabilità alle proteine \u200b\u200bfino a 10 kDa) piegata interna (crista) m-na (75% di proteine): proteine \u200b\u200bdi trasporto, f-you, componenti della catena respiratoria e ATP sintasi, cardiolipina) ( arricchito con ciclo citrato f-tami) inter 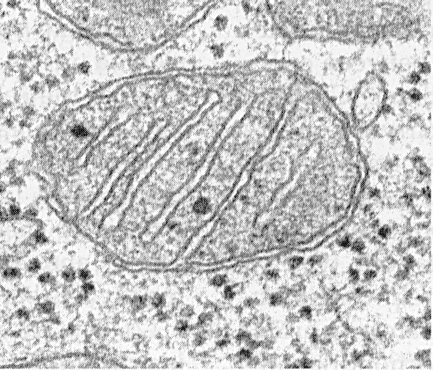
I mitocondri o i condriosomi (dal greco. Mitos - filo, condrione - grano, soma - corpo) sono organelli granulari o filamentosi presenti nel citoplasma di protozoi, piante e animali. I mitocondri possono essere osservati nelle cellule viventi, poiché hanno una densità abbastanza alta. Nelle cellule viventi, i mitocondri possono muoversi, muoversi, fondersi l'uno con l'altro.
In diverse specie, le dimensioni dei mitocondri sono molto variabili, così come la loro forma è variabile (Fig. 199). Tuttavia, nella maggior parte delle cellule, lo spessore di queste strutture è relativamente costante (circa 0,5 micron) e la lunghezza fluttua, raggiungendo fino a 7-60 micron in forme filamentose.
Studiare le dimensioni e il numero di mitocondri non è una cosa così semplice. Ciò è dovuto al fatto che le dimensioni e il numero di mitocondri che sono visibili su sezioni ultrasottili non corrispondono alla realtà.
I calcoli convenzionali mostrano che nella cellula epatica sono presenti circa 200 mitocondri. Ciò rappresenta oltre il 20% del citoplasma totale e circa il 30-35% della quantità totale di proteine \u200b\u200bnella cellula. La superficie di tutti i mitocondri della cellula epatica è 4-5 volte più grande della superficie della sua membrana plasmatica. La maggior parte dei mitocondri si trova negli ovociti (circa 300.000) e nel caos caos dell'ameba gigante (fino a 500.000).
Nelle cellule vegetali verdi, il numero di mitocondri è inferiore rispetto alle cellule animali, poiché parte delle loro funzioni possono essere svolte dai cloroplasti.
La localizzazione dei mitocondri nelle cellule è diversa. Tipicamente, i mitocondri si accumulano vicino a quelle aree del citoplasma in cui è necessaria la formazione di ATP nei mitocondri. Quindi, nel muscolo scheletrico, i mitocondri si trovano vicino alle miofibrille. Negli spermatozoi, i mitocondri formano una cassa a spirale attorno all'asse del flagello; ciò è probabilmente dovuto alla necessità di utilizzare l'ATP per il movimento della coda dello sperma. Allo stesso modo, nei protozoi e in altre cellule dotate di ciglia, i mitocondri sono localizzati direttamente sotto la membrana cellulare alla base delle ciglia, che richiedono l'ATP per funzionare. Negli assoni delle cellule nervose, i mitocondri si trovano vicino alle sinapsi, dove si verifica il processo di trasmissione di un impulso nervoso. Nelle cellule secretorie che sintetizzano grandi quantità di proteine, i mitocondri sono strettamente associati alle zone ergastoplasma; probabilmente forniscono ATP per l'attivazione di aminoacidi e sintesi proteica su ribosomi.
Ultrastruttura dei mitocondri.
I mitocondri, indipendentemente dalle loro dimensioni o forma, hanno una struttura universale, la loro ultrastruttura è uniforme. I mitocondri sono limitati a due membrane (Fig. 205). La membrana mitocondriale esterna la separa dallo ialoplasma, ha contorni lisci, non forma sporgenze o pieghe e lo spessore è di circa 7 nm. Rappresenta circa il 7% dell'area di tutte le membrane cellulari. La membrana non è collegata ad altre membrane citoplasmatiche, è chiusa su se stessa ed è una sacca a membrana. La membrana esterna dall'interno è separata da uno spazio intermembrana con una larghezza di circa 10-20 nm. La membrana interna (circa 7 nm di spessore) limita l'effettivo contenuto interno dei mitocondri, la sua matrice o mitoplasma. La membrana mitocondriale interna forma numerose intrusioni all'interno dei mitocondri. Tali sporgenze il più delle volte hanno la forma di creste piatte o criste.
La superficie totale della membrana mitocondriale interna nella cellula epatica è circa un terzo della superficie di tutte le membrane cellulari. I mitocondri delle cellule del muscolo cardiaco contengono un numero di croste tre volte superiore a quello dei mitocondri epatici, il che riflette le differenze nei carichi funzionali dei mitocondri di cellule diverse. La distanza tra le membrane nella crista è di circa 10-20 nm.
Le malattie mitocondriali che si estendono dalla membrana interna e si estendono verso la matrice non bloccano completamente la cavità mitocondriale e non violano la continuità della matrice che la riempie.
L'orientamento delle creste rispetto all'asse lungo dei mitocondri è diverso per le diverse cellule. L'orientamento può essere creste perpendicolari (fegato, cellule renali); nel muscolo cardiaco, si osserva una disposizione longitudinale di criste. Crista può diramare o formare processi a forma di dito, piegarsi e non avere un orientamento pronunciato (Fig. 208). Nei protozoi, alghe unicellulari, in alcune cellule delle piante e degli animali superiori, le escrescenze della membrana interna sembrano tubi (croste tubulari).
La matrice mitocondriale ha una struttura omogenea a grana fine in cui vengono rilevate molecole di DNA sotto forma di filamenti sottili raccolti in una palla (circa 2-3 nm) e ribosomi mitocondriali sotto forma di granuli di dimensioni circa 15-20 nm. I depositi di magnesio e sali di calcio nella matrice formano granuli densi (20-40 nm).
Le funzioni dei mitocondri.
I mitocondri effettuano la sintesi di ATP, che si verifica a seguito dell'ossidazione dei substrati organici e della fosforilazione dell'ADP.
Le fasi iniziali dell'ossidazione dei carboidrati sono chiamate ossidazione anaerobica, o glicolisi e si verificano nello ialoplasma e non richiedono ossigeno. Il substrato di ossidazione durante la produzione di energia anaerobica sono esosi e principalmente glucosio; alcuni batteri hanno la capacità di estrarre energia ossidando pentosi, acidi grassi o aminoacidi.
Nel glucosio, la quantità di energia potenziale contenuta nei legami tra gli atomi C, H e O è di circa 680 kcal per mole (cioè 180 g di glucosio).
In una cellula vivente, questa enorme quantità di energia viene rilasciata sotto forma di un processo graduale, controllato da un numero di enzimi ossidanti, e non è associato alla transizione dell'energia del legame chimico al calore, come durante la combustione, ma passa in un legame macroenergetico nelle molecole di ATP, che sono sintetizzate usando l'energia rilasciata da ADP e fosfato.
I triosi formati a seguito di glicolisi, principalmente acido piruvico, sono coinvolti in un'ulteriore ossidazione nei mitocondri. In questo caso, si verifica l'uso dell'energia di scissione di tutti i legami chimici, che porta al rilascio di CO 2, al consumo di ossigeno e alla sintesi di una grande quantità di ATP. Questi processi sono associati al ciclo di ossidazione degli acidi tricarbossilici e alla catena respiratoria del trasferimento di elettroni, in cui si verificano la fosforilazione dell'ADP e la sintesi del "combustibile" cellulare e delle molecole di ATP (Fig. 209).
Nel ciclo dell'acido tricarbossilico (il ciclo di Krebs, o ciclo dell'acido citrico), il piruvato formato a causa della glicolisi perde prima la molecola di CO 2 e, essendo ossidato in acetato (un composto a due atomi di carbonio), si combina con il coenzima A. Quindi, il coenzima acetilico A si combina con l'ossalacetato (un composto a quattro atomi di carbonio), forma citrato a sei atomi di carbonio (acido citrico). Quindi c'è un ciclo di ossidazione di questo composto a sei atomi di carbonio a ossalacetato di quattro atomi di carbonio, che si lega nuovamente all'acetil coenzima A e quindi il ciclo si ripete. Durante questa ossidazione vengono rilasciate due molecole di CO 2 e gli elettroni rilasciati durante l'ossidazione vengono trasferiti alle molecole di coenzima dell'accettore (NAD-nicotinamide adenina dinucleotide), che le coinvolgono ulteriormente nella catena di trasferimento degli elettroni. Pertanto, nel ciclo degli acidi tricarbossilici non esiste alcuna sintesi di ATP, ma c'è un'ossidazione delle molecole, il trasferimento di elettroni agli accettori e l'emissione di CO 2. Tutti gli eventi sopra descritti all'interno dei mitocondri si verificano nella loro matrice.
L'ossidazione del substrato iniziale porta al rilascio di CO 2 e acqua, ma l'energia termica non viene rilasciata, come durante la combustione, ma si formano molecole di ATP. Sono sintetizzati da un altro gruppo di proteine \u200b\u200bche non sono direttamente correlate all'ossidazione. Nelle membrane mitocondriali interne, grandi complessi proteici, enzimi, sintesi di ATP si trovano sulla superficie delle membrane che guardano nella matrice. In un microscopio elettronico, sono visibili sotto forma dei cosiddetti corpi "a fungo", che rivestono completamente la superficie delle membrane, guardando nella matrice. I corpi hanno una gamba e una testa, con un diametro di 8-9 nm. Di conseguenza, sia gli enzimi a catena ossidante che gli enzimi di sintesi ATP sono localizzati nelle membrane mitocondriali interne (Fig. 201b).
La catena respiratoria è il sistema principale per convertire l'energia nei mitocondri. Qui, si verifica l'ossidazione sequenziale e il ripristino degli elementi della catena respiratoria, a seguito della quale l'energia viene rilasciata in piccole porzioni. A causa di questa energia, l'ATP è formato da ADP e fosfato in tre punti della catena. Pertanto, dicono che l'ossidazione (trasferimento di elettroni) è associata alla fosforilazione (ADP + Fn → ATP, cioè si verifica la fosforilazione ossidativa.
L'energia rilasciata durante il trasporto di elettroni viene immagazzinata sotto forma di un gradiente di protoni sulla membrana. Si è scoperto che durante il trasferimento di elettroni nella membrana mitocondriale, ogni complesso della catena respiratoria dirige l'energia libera da ossidazione per spostare i protoni (cariche positive) attraverso la membrana, dalla matrice allo spazio intermembrana, che porta alla formazione di potenziali differenze sulla membrana: le cariche positive prevalgono nello spazio intermembrana, e negativo - dal lato della matrice mitocondriale. Quando viene raggiunta la differenza potenziale (220 mV), il complesso proteico della sintetasi di ATP inizia a trasportare i protoni nella matrice, mentre trasforma una forma di energia in un'altra: forma l'ATP dall'ADP e il fosfato inorganico. Questa è la combinazione di processi ossidativi con processi sintetici, con fosforilazione dell'ADP. Mentre i substrati si ossidano, mentre i protoni vengono pompati attraverso la membrana mitocondriale interna, si verifica la sintesi di ATP associata a questo, ad es. si verifica la fosforilazione ossidativa.
Questi due processi possono essere separati. In questo caso, il trasferimento di elettroni continua, così come l'ossidazione del substrato, ma non si verifica la sintesi di ATP. In questo caso, l'energia rilasciata durante l'ossidazione viene convertita in energia termica.
Fosforilazione ossidativa nei batteri.
Nelle cellule procariotiche capaci di fosforilazione ossidativa, gli elementi del ciclo dell'acido tricarbossilico sono localizzati direttamente nel citoplasma e la catena respiratoria e gli enzimi di fosforilazione sono associati alla membrana cellulare, con le sue sporgenze sporgenti all'interno del citoplasma, con i cosiddetti mesosomi (Fig. 212). Va notato che tali mesosomi batterici possono essere associati non solo ai processi di respirazione aerobica, ma anche ad alcune specie partecipano alla divisione cellulare, al processo di distribuzione del DNA in nuove cellule, alla formazione della parete cellulare, ecc.
Sulla membrana plasmatica dei mesosomi di alcuni batteri vengono eseguiti processi coniugati di ossidazione e sintesi di ATP. Particelle sferiche simili a quelle trovate nei mitocondri delle cellule eucariotiche sono state trovate al microscopio elettronico nelle frazioni delle membrane plasmatiche dei batteri. Pertanto, nelle cellule batteriche in grado di fosforilazione ossidativa, la membrana plasmatica svolge un ruolo simile alla membrana mitocondriale interna delle cellule eucariotiche.
Un aumento del numero di mitocondri.
I mitocondri possono aumentare il loro numero soprattutto con la divisione cellulare o con un aumento del carico funzionale della cellula. C'è un costante rinnovamento dei mitocondri. Ad esempio, nel fegato, la durata media dei mitocondri è di circa 10 giorni.
Un aumento del numero di mitocondri si verifica attraverso la crescita e la divisione dei mitocondri precedenti. Questa ipotesi fu formulata per la prima volta da Altman (1893), che descrisse i mitocondri con il termine "bioblasti". È possibile osservare la divisione in vivo, la frammentazione dei mitocondri lunghi in quelli più corti per costrizione, che ricorda il metodo binario di divisione dei batteri.
Un vero aumento del numero di mitocondri per divisione è stato stabilito studiando il comportamento dei mitocondri nelle cellule viventi della coltura dei tessuti. Durante il ciclo cellulare, i mitocondri crescono a diversi micron, e quindi frammentano, si dividono in corpi più piccoli.
I mitocondri possono fondersi l'uno con l'altro e moltiplicarsi secondo il principio: i mitocondri dei mitocondri.
Autoproduzione di mitocondri.
Gli organelli a due membrane hanno un sistema completo di auto-riproduzione. Nei mitocondri e nei plastidi esiste il DNA su cui sono sintetizzati informazioni, trasferimento e RNA ribosomiali e ribosomi, che svolgono la sintesi di proteine \u200b\u200bmitocondriali e plastidiche. Tuttavia, questi sistemi, sebbene autonomi, sono limitati nelle loro capacità.
Il DNA nei mitocondri è una molecola ciclica senza istoni e quindi ricorda i cromosomi batterici. La loro dimensione è di circa 7 micron, 16-19 mila coppie di nucleotidi di DNA entrano in una molecola ciclica di mitocondri animali. Nell'uomo, il DNA mitocondriale contiene 16.5 mila n.p., è completamente decifrato. Si è scoperto che il DNA mitocondrale di vari oggetti è molto omogeneo, la loro differenza sta solo nella grandezza degli introni e dei siti non trascritti. Tutti i DNA mitocondriali sono copie multiple assemblate in gruppi, cluster. Quindi in un mitocondrio del fegato di ratto può contenere da 1 a 50 molecole di DNA ciclico. La quantità totale di DNA mitocondriale per cellula è di circa l'uno percento. La sintesi del DNA mitocondriale non è correlata alla sintesi del DNA nel nucleo.
Come nei batteri, il DNA mitocondrale viene raccolto in una zona separata - il nucleoide, la sua dimensione è di circa 0,4 micron di diametro. Nei mitocondri lunghi possono esserci da 1 a 10 nuclei. Quando si divide un lungo mitocondrio, una sezione contenente un nucleoide viene separata da esso (simile alla divisione binaria dei batteri). La quantità di DNA nei singoli nucleidi mitocondriali può fluttuare 10 volte a seconda del tipo di cellule.
In alcune culture, nelle cellule dal 6 al 60% dei mitocondri non ha un nucleoide, il che può essere spiegato dal fatto che la divisione di questi organelli è più probabilmente associata alla frammentazione e non alla distribuzione dei nuclei.
Come già accennato, i mitocondri possono sia dividersi che fondersi l'uno con l'altro. Quando i mitocondri si fondono tra loro, può verificarsi uno scambio dei loro componenti interni.
È importante sottolineare che rRNA e ribosomi dei mitocondri e del citoplasma sono nettamente diversi. Se nel citoplasma si trovano ribosomi degli anni '80, i ribosomi mitocondriali delle cellule vegetali appartengono ai ribosomi degli anni '70 (sono costituiti da subunità degli anni '30 e '50, contengono gli RNA degli anni '30 e '50 caratteristici delle cellule procariotiche) e i ribosomi più piccoli si trovano nei mitocondri delle cellule animali (circa 50s).
Gli RNA ribosomiali mitocondriali sono sintetizzati sul DNA mitocondriale. Nel mitoplasma sui ribosomi è la sintesi delle proteine. Si ferma, a differenza della sintesi sui ribosomi citoplasmatici, sotto l'azione dell'antibiotico cloramfenicolo, che inibisce la sintesi delle proteine \u200b\u200bnei batteri.
Sul genoma mitocondriale vengono sintetizzati 22 RNA di trasporto. Il codice tripletto del sistema sintetico mitocondriale è diverso da quello utilizzato nell'ialoplasma. Nonostante la presenza apparentemente di tutti i componenti necessari per la sintesi proteica, le piccole molecole di DNA mitocondriale non possono codificare tutte le proteine \u200b\u200bmitocondriali, solo una piccola parte di esse. Quindi DNA di 15 mila n.p. può codificare proteine \u200b\u200bcon un peso molecolare totale di circa 6x10 5. Allo stesso tempo, il peso molecolare totale delle proteine \u200b\u200bdelle particelle dell'insieme respiratorio completo dei mitocondri raggiunge un valore di circa 2x10 6.
Dato che oltre alle proteine \u200b\u200bossidative della fosforilazione, i mitocondri includono anche enzimi del ciclo dell'acido tricarbossilico, enzimi di sintesi del DNA e dell'RNA, enzimi di attivazione degli aminoacidi e altre proteine, per codificare queste numerose proteine \u200b\u200be rRNA e tRNA, la quantità di informazioni genetiche contenute la molecola di DNA mitocondriale corta non è chiaramente sufficiente. La decodifica della sequenza nucleotidica del DNA mitocondriale umano ha dimostrato che codifica solo 2 RNA ribosomiali, 22 RNA di trasferimento e un totale di 13 diverse catene polipeptidiche.
È stato ora dimostrato che la maggior parte delle proteine \u200b\u200bmitocondriali sono sotto il controllo genetico dal nucleo cellulare e sono sintetizzate al di fuori dei mitocondri. La maggior parte delle proteine \u200b\u200bmitocondriali sono sintetizzate sui ribosomi nel citosol. Queste proteine \u200b\u200bhanno sequenze di segnali speciali che sono riconosciute dai recettori sulla membrana mitocondriale esterna. Queste proteine \u200b\u200bpossono essere inserite in esse (vedere l'analogia con la membrana perossisomica), quindi spostarsi sulla membrana interna. Questo trasferimento avviene nei punti di contatto delle membrane esterne ed interne in cui si nota tale trasporto. La maggior parte dei lipidi mitocondriali sono anche sintetizzati nel citoplasma.
Tutto ciò indica l'origine endosimbiotica dei mitocondri, che i mitocondri sono organismi come i batteri, che sono in simbiosi con una cellula eucariotica.
Condrite.
La totalità di tutti i mitocondri in una cellula è chiamata condrioma. Può essere diverso a seconda del tipo di celle. In molte cellule, i condriomi sono costituiti da numerosi mitocondri sparsi situati uniformemente in tutto il citoplasma o localizzati in gruppi nei siti di intenso dispendio di ATP. In entrambi questi casi, i mitocondri funzionano da soli, il loro lavoro cooperativo, eventualmente coordinato da alcuni segnali del citoplasma. Esiste un tipo completamente diverso di condrioma, quando invece di piccoli mitocondri sparsi singoli, nella cellula si trova un mitocondrio ramificato gigante.
Tali mitocondri si trovano nelle alghe verdi unicellulari (ad esempio la clorella). Formano una complessa rete mitocondriale o reticolo mitocondriale (Reticulum miyochondriale). Secondo la teoria chemio osmotica, il significato biologico dell'apparizione di una struttura mitocondriale ramificata così gigante, unita dalle sue membrane esterne ed interne, è che la sintesi di ATP può avvenire in qualsiasi punto sulla superficie della membrana interna di tali mitocondri ramificati, che andrà ovunque nel citoplasma, dove C'è bisogno di questo.
Nel caso dei mitocondri ramificati giganti, in qualsiasi punto della membrana interna, può accumularsi un potenziale sufficiente per iniziare la sintesi di ATP. Da queste posizioni, il reticolo mitocondriale è, per così dire, un conduttore elettrico, un cavo che collega i punti distanti di un tale sistema. Il reticolo mitocondriale si è rivelato molto utile non solo per le piccole cellule mobili, come la clorella, ma anche per unità strutturali più grandi come, ad esempio, miofibrille nel muscolo scheletrico.
È noto che il muscolo scheletrico è costituito da una massa di fibre muscolari, simbologie contenenti molti nuclei. La lunghezza di tali fibre muscolari raggiunge i 40 micron, con uno spessore di 0,1 micron: questa è una struttura gigantesca contenente moltissime miofibrille, che si contraggono simultaneamente, in modo sincrono. Per la contrazione, una grande quantità di ATP, che i mitocondri forniscono a livello di z-disk, viene erogata a ciascuna unità di contrazione, a miofibrilla. Sulle sezioni ultrasottili longitudinali dei muscoli scheletrici al microscopio elettronico, sono visibili numerose piccole sezioni arrotondate di mitocondri situati in prossimità dei sarcomeri. I mitocondri muscolari non sono piccole sfere o bastoncini, ma piuttosto strutture aracnidi, i cui processi si ramificano e si estendono su lunghe distanze, a volte attraverso l'intero diametro della fibra muscolare.
Allo stesso tempo, la ramificazione mitocondriale circonda ogni miofibrilla nella fibra muscolare, fornendo loro l'ATP necessario per la contrazione muscolare. Pertanto, nel piano del disco z, i mitocondri rappresentano un tipico reticolo mitocondriale. Tale strato o piano del reticolo mitocondriale viene ripetuto due volte per ciascun sarcomero e l'intera fibra muscolare ha migliaia di strati trasversali di "pavimento" del reticolo mitocondriale. Tra i "piani" lungo le miofibrille vi sono mitocondri filamentosi che collegano questi strati mitocondriali. Pertanto, è stata creata un'immagine tridimensionale del reticolo mitocondriale, che passa attraverso l'intero volume della fibra muscolare.
È stato inoltre stabilito che tra i rami del reticolo mitocondriale e i mitocondri longitudinali filiformi ci sono composti o contatti intermitocondriali speciali (MMC). Sono formati da membrane mitocondriali esterne strettamente adiacenti di mitocondri a contatto, lo spazio intermembrana e le membrane in questa zona hanno una densità di elettroni aumentata. Attraverso queste formazioni speciali, l'unione funzionale dei mitocondri vicini e dei reticoli mitocondriali avviene in un unico sistema energetico cooperativo. Tutte le miofibrille nella fibra muscolare sono ridotte in modo sincrono per tutta la loro lunghezza, quindi la fornitura di ATP su qualsiasi parte di questa complessa macchina dovrebbe avvenire in modo sincrono, e ciò può avvenire solo se un numero enorme di conduttori mitocondri ramificati sono collegati tra loro usando i contatti.
Il fatto che i contatti intermitocodiali (MMK) siano coinvolti nell'associazione energetica dei mitocondri tra loro era possibile su cardiomiociti, cellule muscolari del cuore.
I condriomi delle cellule muscolari del cuore non formano strutture ramificate, ma sono rappresentati da molti piccoli mitocondri allungati situati senza ordine speciale tra le miofibrille. Tuttavia, tutti i mitocondri vicini si uniscono tra loro usando contatti mitocondriali dello stesso tipo dei muscoli scheletrici, solo il loro numero è molto grande: in media, ci sono 2-3 MMC per mitocondri che legano i mitocondri in una singola catena, dove ogni anello ha la catena (Streptio mitochondriale) è un mitocondrio separato.
Si è scoperto che i contatti intermitocondriali (MMC), come struttura obbligatoria delle cellule cardiache, sono stati trovati nei cardiomiociti sia dei ventricoli che degli atri di tutti i vertebrati: mammiferi, uccelli, rettili, anfibi e pesci ossei. Inoltre, sono stati trovati MMC (ma in numero minore) nelle cellule cardiache di alcuni insetti e molluschi.
La quantità di MMC nei cardiomiociti varia a seconda del carico funzionale sul cuore. Il numero di MMC aumenta con l'aumentare dell'attività fisica degli animali e, al contrario, quando il carico sul muscolo cardiaco diminuisce, si verifica una forte riduzione del numero di MMC.







