Comme les structures internes des mitochondries sont appelées. La structure et les fonctions des mitochondries et des plastes
Mitochondries
Dans les cellules des tissus animaux, des mitochondries ont été trouvées en 1882, et dans les plantes seulement en 1904 (dans les anthères de nénuphars). Il a été possible d'établir des fonctions biologiques après séparation et purification de la fraction par centrifugation fractionnée. Ils contiennent 70% de protéines et environ 30% de lipides, une petite quantité d'ARN et d'ADN, des vitamines A, B 6, B 12, K, E, des acides folique et pantothénique, de la riboflavine, diverses enzymes. Les mitochondries ont une double membrane, l'extérieur isole l'organite du cytoplasme et l'intérieur forme des excroissances de crista. Tout l'espace entre les membranes est rempli d'une matrice (Fig. 13).
La fonction principale des mitochondries est la participation à la respiration cellulaire. Le rôle des mitochondries dans la respiration a été établi en 1950-1951. Un système enzymatique complexe du cycle de Krebs est concentré sur les membranes externes. Pendant l'oxydation des substrats respiratoires, de l'énergie est libérée qui est immédiatement accumulée dans les molécules d'ADP formées, principalement l'ATP, pendant la phosphorylation oxydative qui se produit dans les crêtes. L'énergie stockée dans les composés macroergiques est utilisée à l'avenir pour répondre à tous les besoins de la cellule.
La formation de mitochondries dans la cellule se produit en continu à partir des micro-organismes, le plus souvent leur apparition est associée à la différenciation des structures membranaires de la cellule. Ils peuvent être restaurés dans la cellule en les divisant et en les bourgeonnant. Les mitochondries ne sont pas durables, leur espérance de vie est de 5 à 10 jours.
Les mitochondries sont des stations cellulaires «électriques». Ils concentrent l'énergie, qui est stockée dans les «accumulateurs» d'énergie - les molécules d'ATP, et non dissipée dans la cellule. La violation de la structure des mitochondries conduit à une perturbation du processus respiratoire et, par conséquent, à la pathologie du corps.
Appareil de Golgi.Appareil de Golgi (un synonyme - dictiosomes) est un empilement de 3-12 aplatis, fermés, entourés d'une double membrane de disques appelés réservoirs, à partir des bords desquels de nombreuses bulles sont décollées (300-500). La largeur des réservoirs est de 6-90 A, l'épaisseur des membranes est de 60-70 A.
L'appareil de Golgi est un centre de synthèse, d'accumulation et d'isolement de polysaccharides, en particulier de cellulose, et est impliqué dans la distribution et le transport intracellulaire des protéines, ainsi que dans la formation de vacuoles et de lysosomes. Dans la cellule végétale, il a été possible de retracer la participation de l'appareil de Golgi à l'apparition de la plaque médiane et la croissance de la membrane pecto-cellulosique cellulaire.
L'appareil de Golgi est le plus développé pendant la période de vie cellulaire active. Avec son vieillissement, il s'atrophie progressivement, puis disparaît.
Lysosomes.Lysosomes - corps arrondis plutôt petits (environ 0,5 microns de diamètre). Ils sont recouverts d'une membrane protéique-lipoïde. Le contenu des lysosomes est constitué de nombreuses enzymes hydrolytiques qui remplissent la fonction de digestion intracellulaire (lyse) des macromolécules protéiques, des acides nucléiques, des polysaccharides. Leur fonction principale est la digestion de sections individuelles du protoplaste cellulaire (autophagie - auto-dévorante). Ce processus se déroule en raison d'une phagocytose ou d'une pinocytose. Le rôle biologique de ce processus est double. Premièrement, elle est protectrice, car avec une pénurie temporaire de produits de rechange, la cellule soutient la vie en raison de protéines constitutionnelles et d'autres substances, et deuxièmement, il y a une libération d'organes en excès ou usés (plastes, mitochondries, etc.) La membrane du lysosome empêche la libération d'enzymes dans le cytoplasme sinon, il serait complètement digéré par ces enzymes.
Dans une cellule morte, les lysosomes sont détruits, les enzymes sont dans la cellule et tout son contenu est digéré. Il ne reste que la coque en pecto-cellulose.
Les lysosomes sont des produits de l'activité de l'appareil de Golgi, des vésicules qui s'en détachent, dans lesquelles cet organoïde accumule des enzymes de digestion.
Sphérosomes - corps protéines-lipoïdes arrondis 0,3-0,4 microns. Selon toute vraisemblance, ce sont des dérivés de l'appareil de Golgi ou du réticulum endoplasmique. Dans leur forme et leur taille ressemblent à des lysosomes. Étant donné que les sphérosomes contiennent de la phosphatase acide, ils sont probablement liés aux lysosomes. Certains auteurs pensent que les sphérosomes et les lysosomes sont équivalents les uns aux autres, mais très probablement uniquement en termes d'origine et de forme. Il y a une hypothèse sur leur participation à la synthèse des graisses (A. Frey-Wissling).
Ribosomes - de très petits organoïdes, leur diamètre est d'environ 250 A. De forme presque sphérique. Certains d'entre eux sont attachés aux membranes externes du réticulum endoplasmique, certains sont à l'état libre dans le cytoplasme. Une cellule peut contenir jusqu'à 5 millions de ribosomes. Les ribosomes se trouvent dans les chloroplastes et les mitochondries, où ils synthétisent une partie des protéines dont ces organoïdes sont construits et les enzymes qui y fonctionnent.
La fonction principale est la synthèse de protéines spécifiques en fonction des informations provenant du noyau. Leur composition: protéine et acide ribonucléique ribosomique (ARN) en proportions égales. Leur structure est constituée de petites et grandes sous-unités formées de ribonucléotide.
Microtubules.Microtubules - dérivés particuliers du réticulum endoplasmique. Trouvé dans de nombreuses cellules. Leur nom lui-même parle de leur forme - un ou deux, situés en parallèle, des tubules avec une cavité à l'intérieur. Diamètre extérieur dans 250A. Les parois des microtubules sont construites à partir de molécules de protéines. Les filaments du fuseau se forment à partir des microtubules lors de la division cellulaire.
Le noyau
Le noyau a été découvert dans une cellule végétale par R. Brown en 1831. Il est situé au centre de la cellule ou près de la membrane cellulaire, mais est entouré de cytoplasme de tous les côtés. Dans la plupart des cas, il y a un noyau dans la cellule, plusieurs noyaux se trouvent dans les cellules de certaines algues, ainsi que des champignons. Les algues vertes non cellulaires ont des centaines de noyaux. Cellules multinucléées de bactéries non articulées. Il n'y a pas de noyaux dans les cellules des bactéries et des algues bleu-vert.
La forme du noyau est le plus souvent proche de la forme d'une boule ou d'une ellipse. Dépend de la forme, de l'âge et de la fonction de la cellule. Dans la cellule méristématique, le noyau est gros, de forme ronde et occupe les 3/4 du volume cellulaire. Dans les cellules parenchymateuses de l'épiderme ayant une grande vacuole centrale, le noyau a une forme lenticulaire et est poussé avec le cytoplasme à la périphérie de la cellule. C'est le signe d'une cellule spécialisée, mais déjà vieillissante. Une cellule dépourvue de noyau ne peut vivre que peu de temps. Les cellules exemptes d'armes nucléaires des tubes de tamis sont des cellules vivantes, mais elles ne vivent pas longtemps. Dans tous les autres cas, les cellules exemptes d'armes nucléaires sont mortes.
Le noyau a une double coque, à travers les pores dans lesquels le contenu
les noyaux (nucléoplasme) peuvent communiquer avec le contenu du cytoplasme. Les membranes membranaires du noyau sont équipées de ribosomes et communiquent avec les membranes du réticulum endoplasmique de la cellule. Un ou deux nucléoles et chromosomes sont situés dans le nucléoplasme. Le nucléoplasme est un système colloïdal de Zola, qui rappelle une gélatine épaissie de consistance. Selon les biochimistes russes (I.Zbarsky, etc.), le noyau contient quatre fractions de protéines: protéines simples - globulines 20%, désoxyribonucléoprotéines - 70%, protéines acides - 6% et protéines résiduelles 4%. Ils sont localisés dans les structures nucléaires suivantes: ADN protéides (protéines alcalines) dans les chromosomes, ARN protéides (protéines acides) dans les nucléoles, en partie dans les chromosomes (lors de la synthèse de l'ARN messager) et dans la membrane nucléaire. Les globulines constituent la base du nucléoplasme. Les protéines résiduelles (nature non précisée) forment une membrane nucléaire.
La majeure partie des protéines de base sont des protéines alcalines complexes de désoxyribonucléoprotéines, qui sont basées sur l'ADN.
Molécule d'ADN.Molécule d'ADN - polynucléotide et se compose de nucléotides. Le nucléotide contient trois composants: une molécule de sucre (désoxyribose), une molécule azotée et une molécule d'acide phosphorique. Le désoxyribose est lié à la base glycosidique azotée et à l'acide phosphorique - une liaison éther. Dans l'ADN, dans diverses combinaisons, il n'y a que 4 variétés de nucléotides qui diffèrent les unes des autres par leurs bases azotées. Deux d'entre eux (adénine et guanine) appartiennent à des composés azotés puriques, et la cytosine et la thymine à la pyrimidine. Les molécules d'ADN ne sont pas situées sur le même plan, mais se composent de deux brins en spirale, c'est-à-dire deux chaînes parallèles torsadées l'une autour de l'autre forment une molécule d'ADN. Ils sont liés les uns aux autres via une liaison hydrogène entre les bases azotées, et les bases purines d'une chaîne fixent les bases pyrimidiques de l'autre (Fig. 14). La structure et la chimie de la molécule d'ADN ont été découvertes par des scientifiques anglais (Crick) et américains (Watson) et publiées en 1953. Ce point est considéré comme le début du développement de la génétique moléculaire. Le poids moléculaire de l'ADN est de 4 à 8 millions et le nombre de nucléotides (diverses options) peut atteindre 100 000. La molécule d'ADN est très stable, sa stabilité est assurée par le fait qu'à travers elle, elle a la même épaisseur - 20A (8A - la largeur de la base pyrimidine + 12A - la largeur de la base purine). Si du phosphore radioactif est introduit dans l'organisme, le marqueur sera détecté dans tous les composés contenant du phosphore, à l'exception de l'ADN (Levy, Sickewitz).
Les molécules d'ADN sont porteuses d'hérédité, car leur structure code des informations sur la synthèse de protéines spécifiques qui déterminent les propriétés de l'organisme. Des changements peuvent se produire sous l'influence de facteurs mutagènes (rayonnement radioactif, agents chimiques puissants, alcaloïdes, alcools, etc.).
Molécule d'ARN.Molécules d'acide ribonucléique (ARN) beaucoup moins de molécules d'ADN. Ce sont des chaînes uniques de nucléotides. Il existe trois types d'ARN: le ribosomal, le plus long, formant de nombreuses boucles, l'information (matrice) et le transport, le plus court. L'ARN ribosomal est localisé dans les ribosomes du réticulum endoplasmique et représente 85% de l'ARN total de la cellule.
L'ARN informationnel dans sa structure ressemble à une feuille de trèfle. Sa quantité est de 5% de l'ARN total dans la cellule. Il est synthétisé dans les nucléoles. Son assemblage se produit dans les chromosomes pendant la période d'interphase. Sa fonction principale est le transfert d'informations de l'ADN aux ribosomes, où la synthèse des protéines a lieu.
L'ARN de transport, tel qu'il est maintenant établi, est une famille entière de composés liés par leur structure et leur fonction biologique. Chaque cellule vivante, selon une estimation approximative, contient 40 à 50 ARN de transport individuels et leur nombre total dans la nature, étant donné les différences d'espèces, est énorme. (Acad. V. Engelhardt). Ils sont appelés transport parce que leurs molécules participent au maintien du transport du processus intracellulaire de synthèse des protéines. Combinés avec des acides aminés libres, ils les livrent aux ribosomes de la chaîne protéique en construction. Ce sont les plus petites molécules d'ARN, composées en moyenne de 80 nucléotides. Localisé dans la matrice du cytoplasme et représente environ 10% de l'ARN cellulaire
L'ARN contient quatre bases azotées, mais contrairement à l'ADN, l'uracile remplace la thymine au lieu de la thymine.
La structure des chromosomes.Les chromosomes ont été découverts pour la première fois à la fin du 19e siècle par les classiques de la cytologie Fleming et Strasburger (1882, 1884), et le chercheur russe I.D. Chistyakov les a découverts en 1874.
L'élément structurel principal des chromosomes est le noyau. Ils ont une forme différente. Ce sont des bâtons droits ou courbes, des corps ovales, des boules, dont les tailles varient.
Selon l'emplacement du centromère, les chromosomes directs, égaux et inégaux sont distingués. La structure interne des chromosomes est représentée sur la Fig. 15, 16. Il convient de noter que la désoxyribonucléoprotéine est un monomère du chromosome.
Dans le chromosome des désoxyribonucléoprotéines, 90-92%, dont 45% d'ADN et 55% de protéines (histone). En petite quantité dans le chromosome, l'ARN est également représenté (informationnel).
Le chromosome a également une structure transversale prononcée - la présence de zones épaissies - des disques, qui étaient de retour en 1909. ont été appelés gènes. Ce terme a été proposé par le scientifique danois Johansen. En 1911, le scientifique américain Morgan a prouvé que les gènes sont les principales unités héréditaires et qu'ils sont répartis dans les chromosomes dans un ordre linéaire et, par conséquent, le chromosome a des sections qualitativement différentes. En 1934, le scientifique américain Pinter a prouvé la discontinuité de la structure morphologique des chromosomes et la présence de disques dans les chromosomes, et les disques sont des endroits où l'ADN s'accumule. Cela a servi de début à la création de cartes chromosomiques, qui indiquaient l'emplacement (locus) de l'emplacement du gène qui détermine l'un ou l'autre des traits de l'organisme. Un gène est un site d'ADN à double hélice qui contient des informations sur la structure d'une seule protéine. Il s'agit d'une section d'une molécule d'ADN qui détermine la synthèse d'une seule molécule de protéine. L'ADN ne participe pas directement à la synthèse des protéines. Il contient et stocke uniquement des informations sur la structure de la protéine.
La structure de l'ADN, composée de plusieurs milliers de 4 nucléotides situés consécutivement, est un code d'hérédité.
Le code d'hérédité. Synthèse des protéines.Le premier message sur le code ADN a été fait par le biochimiste américain Nirenberg en 1961 à Moscou lors du Congrès international de biochimie. L'essence du code ADN est la suivante. Chaque acide aminé correspond à une partie de la chaîne d'ADN de trois nucléotides adjacents (triplets). Ainsi, par exemple, un site constitué de TT-T (un triplet de 3 nucléotides contenant de la thymine) correspond à l'acide aminé lysine, un triplet A (adénine) - C (cytosine) - A (adénine) - cystéine, etc. Supposons qu'un gène soit représenté par une chaîne de nucléotides située dans l'ordre suivant: A-C-A-T-T-T-A-A-C-C-A-A-G-G-G. Après avoir divisé cette série en triplets, nous allons immédiatement déchiffrer quels acides aminés et dans quel ordre seront situés dans la protéine synthétisée.
Le nombre de combinaisons possibles de 4 nucléotides disponibles de trois est 4 × 64. Sur la base de ces ratios, le nombre de triplets différents en excès sera suffisant pour fournir des informations sur la synthèse de nombreuses protéines qui déterminent à la fois la structure et les fonctions du corps. Pour la synthèse des protéines, une copie exacte de ces informations est envoyée aux ribosomes sous forme d'ARN messager. Dans le décodage et la synthèse, en plus de l'i-ARN, un grand nombre de molécules de divers acides ribonucléiques de transport (ARN-t), de ribosomes et d'un certain nombre d'enzymes sont impliquées. Chacun des 20 acides aminés se lie à l'ARN-T - une molécule avec une molécule. Chacun des 20 acides aminés a son propre ARN-t. Les ARN-T ont des groupes chimiques capables de «reconnaître» leur acide aminé, en le choisissant parmi les acides aminés disponibles. Cela se produit à l'aide d'enzymes spéciales. Ayant reconnu son acide aminé, l'ARN-t entre dans un composé avec lui. Un ribosome est attaché au début de la chaîne (molécule) de l'i-ARN qui, se déplaçant le long de l'i-ARN, se connecte les uns aux autres dans la chaîne polypeptidique exactement les acides aminés dont l'ordre est codé par la séquence nucléotidique de cet I-ARN. En conséquence, une molécule de protéine est formée, dont la composition est codée dans l'un des gènes.
Nucleoli- une partie structurelle intégrale du noyau. Ce sont des corps sphériques. Ils sont très variables, changent de forme et de structure, apparaissent et disparaissent. Il y en a un, deux. Pour chaque plante un certain nombre. Les noyaux disparaissent lorsque la cellule se prépare à la division, puis réapparaissent; ils sont apparemment impliqués dans la synthèse des acides ribonucléiques. Si le nucléole est détruit par un faisceau focalisé de rayons X ou ultraviolets, la division cellulaire est supprimée.
Le rôle du noyau dans la vie cellulaire. Le noyau sert de centre de contrôle de la cellule - il dirige l'activité cellulaire et contient des porteurs d'hérédité (gènes) qui déterminent les caractéristiques d'un organisme donné. Le rôle du noyau peut être identifié si, à l'aide de techniques microchirurgicales, le retirer de la cellule et en observer la conséquence. Une série d'expériences prouvant un rôle important dans la régulation de la croissance cellulaire a été réalisée par Hemmerling sur l'algue verte unicellulaire Acetobularia. Cette algue atteint une hauteur de 5 cm, ressemble à un champignon, a la ressemblance de «racines» et de «pattes». Au-dessus se termine par un grand "chapeau" en forme de disque. La cellule de cette algue possède un noyau situé dans la partie basale de la cellule.
Hammerling a constaté que si la jambe est coupée, la partie inférieure continue de vivre et le capuchon est complètement régénéré après l'opération. La partie supérieure, dépourvue de noyau, survit un certain temps, mais finit par mourir, ne pouvant restaurer la partie inférieure. Par conséquent, le noyau d'acétobularia est nécessaire pour les réactions métaboliques sous-jacentes à la croissance.
Le noyau favorise la formation de la paroi cellulaire. Cela peut être illustré par des expériences avec l'algue Voucheria et Spyrogyra. En libérant le contenu des cellules des filaments coupés dans l'eau, nous pouvons obtenir des morceaux de cytoplasme avec un, avec plusieurs noyaux et sans noyaux. Dans les deux premiers cas, la membrane cellulaire s'est formée normalement. En l'absence de noyau, la coque ne s'est pas formée.
Dans les expériences de I.I. Gerasimov (1890) avec le spirogyre, il a été constaté que les cellules à double noyau doublent la longueur et l'épaisseur du chloroplaste. Dans les cellules non nucléaires, le processus de photosynthèse se poursuit, l'amidon d'assimilation se forme, mais le processus de son hydrolyse est atténué, ce qui s'explique par l'absence d'enzymes hydrolytiques qui ne peuvent être synthétisées dans les ribosomes qu'en fonction de l'ADN du noyau. La durée de vie d'un protoplaste sans noyau est inférieure et de courte durée. Dans les expériences de I.I. Les cellules spirogyres exemptes d’énergie nucléaire de Gerasimov ont vécu 42 jours et sont mortes. L'une des fonctions les plus importantes du noyau est d'alimenter le cytoplasme en acide ribonucléique, nécessaire à la synthèse des protéines dans la cellule. L'élimination du noyau de la cellule entraîne une diminution progressive de la teneur en ARN dans le cytoplasme et un ralentissement de la synthèse des protéines en son sein.
Le rôle le plus important du noyau est dans le transfert de traits d'une cellule à l'autre, d'un organisme à un autre, et il le fait dans le processus de fission nucléaire et la cellule dans son ensemble.
Division cellulaire.Les cellules se multiplient par division. Dans ce cas, deux cellules filles avec le même ensemble de matériel héréditaire enfermé dans des chromosomes que la cellule mère sont formées à partir d'une cellule. Dans les cellules somatiques, les chromosomes sont représentés par deux, les chromosomes dits homologues, dans lesquels les gènes alléliques (porteurs de traits opposés, par exemple, les pétales de pois blancs et rouges, etc.), les traits de deux paires parentales sont intégrés. À cet égard, dans les cellules somatiques du corps végétal, il y a toujours un double ensemble de chromoses, noté 2p. Les chromosomes ont une personnalité prononcée. La quantité et la qualité des chromosomes sont une caractéristique de chaque espèce. Ainsi, dans les cellules de fraises, l'ensemble chromoïde diploïde est de 14, (2n), le pommier est de 34, l'artichaut de Jérusalem est de 102, etc.
Mitose (caryocinèse) - la division cellulaire somatique a été décrite pour la première fois par E. Russov (1872) et I.D. Chistyakov (1874). Son essence réside dans le fait que deux cellules filles avec le même ensemble de chromoses sont formées à partir de la cellule mère par division.Le cycle cellulaire est composé d'interphase et de mitose proprement dite. La méthode de microautoradiographie a révélé que la plus longue et la plus difficile est l'interphase - la période du noyau "au repos", car au cours de cette période, il y a un doublement des matières nucléaires. L'interphase est divisée en trois phases:
Q1 - présynthétique (sa durée est de 4 à 6 heures);
S - synthétique (10-20 heures);
Q2 - post-synthétique (2-5 heures).
Pendant la phase Q1, les préparatifs sont en cours pour la reduplication de l'ADN. Et dans la phase S, l'ADN est réduit, la cellule double l'apport d'ADN. Dans la phase Q2, les enzymes et les structures nécessaires pour déclencher la mitose se forment. Ainsi, en interphase, les molécules d'ADN dans les chromosomes sont divisées en deux brins identiques, et l'ARN d'information est assemblé sur leur matrice. Ce dernier porte des informations sur la structure de protéines spécifiques dans le cytoplasme, et dans le noyau, chacun des brins d'ADN complète la moitié manquante de sa molécule. Dans ce processus de doublement (réduction), une caractéristique unique de l'ADN se manifeste, qui consiste en la capacité de l'ADN de se reproduire avec précision. Les molécules d'ADN fille résultantes sont automatiquement obtenues par des copies exactes de la molécule parente, car lors de la reduplication, des bases complémentaires (АТ; Г-Ц; etc.) de l'environnement rejoignent chaque moitié.
Dans la prophase de la division mitotique, des chromosomes doublés deviennent perceptibles. En métaphase, ils sont tous situés dans la zone équatoriale, située dans une rangée. Des filetages de broche sont formés (à partir de microtubules se connectant les uns aux autres). La coquille du noyau et du nucléole disparaissent. Les chromosomes épaissis sont divisés longitudinalement en deux chromosomes filles. C'est l'essence de la mitose. Il fournit une distribution précise des molécules d'ADN doublées entre les cellules filles. Cela garantit la transmission d'informations héréditaires cryptées dans l'ADN.
En anaphase, les chromosomes filles commencent à se déplacer vers des pôles opposés. Au centre, les premiers fragments de la membrane cellulaire (fragmoblaste) apparaissent.
Dans la télophase, les noyaux se forment dans les cellules filles. Le contenu de la cellule mère (organelle) est réparti entre les cellules filles résultantes. La membrane cellulaire est complètement formée. Cela met fin à la cytokinèse (Fig. 17).
Méiose - Division de la réduction a été découvert et décrit dans les années 90 du siècle dernier par V.I.Belyaev. L'essence de la division est qu'à partir d'une cellule somatique contenant un ensemble de chromoses 2n (double, diploïde), quatre cellules haploïdes sont formées, avec "n", un demi-ensemble de chromoses. Ce type de division est complexe et comprend deux étapes. Le premier est la réduction des chromosomes. Les chromosomes doublés sont situés dans la zone équatoriale par paires (deux chromosomes homologues parallèles). À ce moment, une conjugaison (adhésion) par chromose, un croisement (croisement) peut se produire et, par conséquent, un échange de sites de chromose se produit. À la suite de cela, une partie des gènes des chromosomes paternels devient une partie des chromoses maternelles et vice versa. L'apparence des deux chromosomes à la suite de cela ne change pas, mais leur composition qualitative devient différente. Les hérédités paternelles et maternelles sont redistribuées et mixtes.
Dans l'anaphase de la méiose, les chromosomes homologues à l'aide de filaments de fuseau divergent aux pôles, au cours desquels, après une courte période de repos (les filaments disparaissent, mais le septum entre les nouveaux noyaux ne se forme pas), le processus de mitose commence - une métaphase dans laquelle tous les chromosomes sont situés dans un plan et ils sont divisés longitudinalement sur les chromosomes filles. Avec l'anaphase de la mitose à l'aide d'un fuseau, ils divergent aux pôles, où se forment quatre noyaux et, par conséquent, quatre cellules haploïdes. Dans les cellules de certains tissus au cours de leur développement sous l'influence de certains facteurs, une mitose incomplète se produit et le nombre de chromoses dans les noyaux double du fait qu'elles ne divergent pas aux pôles. À la suite de telles violations de nature naturelle ou artificielle, des tétraploïdes et des polyploïdes surviennent. Avec l'aide de la méiose, des cellules germinales se forment - des gamètes, ainsi que des spores, éléments de la reproduction sexuelle et asexuée des plantes (Fig.18).
L'amitose est une fission directe du noyau. Avec l'amitose, un fuseau de division ne se forme pas et la coquille du noyau ne se décompose pas, comme dans la mitose. Auparavant, l'amitose était considérée comme une forme primitive de division. Il est maintenant établi qu'il est associé à la dégradation du corps. Il s'agit d'une version simplifiée d'une fission plus complexe du noyau. L'amitose se produit dans les cellules et les tissus du nucelle, de l'endosperme, du parenchyme tuberculeux, des pétioles des feuilles, etc.
Mitochondries - Il s'agit d'un organite spiralé, rond, allongé ou ramifié.
Le concept de mitochondries a été proposé pour la première fois par Benda en 1897. Les mitochondries peuvent être détectées dans les cellules vivantes en utilisant le contraste de phase et la microscopie d'interférence sous forme de grains, de granules ou de filaments. Ce sont des structures assez mobiles qui peuvent se déplacer, se fusionner, se partager. Lorsqu'elles sont colorées avec des méthodes spéciales dans des cellules mortes sous microscopie optique, les mitochondries ressemblent à de petits grains (granules) qui sont distribués de manière diffuse dans le cytoplasme ou concentrés dans certaines zones spécifiques.
À la suite de la destruction du glucose et des graisses en présence d'oxygène, de l'énergie est générée dans les mitochondries et la matière organique est convertie en eau et en dioxyde de carbone. De cette façon, les organismes animaux reçoivent l'énergie de base nécessaire à la vie. L'énergie s'accumule dans l'adénosine triphosphate (ATP) et plus précisément dans ses liaisons macroergiques. La fonction des mitochondries est étroitement liée à l'oxydation des composés organiques et à l'utilisation de l'énergie libérée lors de leur désintégration pour la synthèse des molécules d'ATP. Par conséquent, les mitochondries sont souvent appelées stations d'énergie cellulaire ou organites de la respiration cellulaire. L'ATP agit en tant que fournisseur d'énergie, transférant l'un de ses groupes phosphates terminaux riches en énergie vers une autre molécule, et se transforme en ADP.
On pense que dans l'évolution, les mitochondries étaient des micro-organismes procaryotes qui sont devenus des symbiotiques dans le corps d'une cellule ancienne. Par la suite, ils sont devenus d'une nécessité vitale, ce qui a été associé à une augmentation de la teneur en oxygène dans l'atmosphère terrestre. D'une part, les mitochondries ont éliminé l'excès d'oxygène toxique pour la cellule et, d'autre part, elles ont fourni de l'énergie.
Sans mitochondries, la cellule est pratiquement incapable d'utiliser l'oxygène comme substance fournissant de l'énergie et ne peut répondre à ses besoins énergétiques que par des processus anaérobies. Ainsi, l'oxygène est un poison, mais le poison est vital pour la cellule, et un excès d'oxygène est tout aussi nocif que sa carence.
Les mitochondries peuvent changer de forme et se déplacer vers les zones de la cellule où elles en ont le plus besoin. Ainsi, dans les cardiomyocytes, les mitochondries sont situées près des myofibrilles, dans les cellules des tubules rénaux près des saillies basales, etc. La cellule contient jusqu'à mille mitochondries, et leur nombre dépend de l'activité de la cellule.
Les mitochondries ont une taille transversale moyenne de 0,5 ... 3 microns. Selon la taille, les mitochondries petites, moyennes, grandes et géantes sont distinguées (elles forment un réseau ramifié - le réticulum mitochondrial). La taille et le nombre de mitochondries sont étroitement liés à l'activité de la cellule et à sa consommation d'énergie. Ils sont extrêmement variables et, en fonction de l'activité cellulaire, de la teneur en oxygène, des influences hormonales, peuvent gonfler, changer le nombre et la structure des crêtes, varier en nombre, forme et taille, ainsi que l'activité enzymatique.
La densité apparente des mitochondries, le degré de développement de leur surface interne et d'autres indicateurs dépendent des besoins énergétiques de la cellule. Dans les lymphocytes, il n'y a que quelques mitochondries, et dans les cellules hépatiques, il y en a 2 ... 3 000.
Les mitochondries sont constituées d'une matrice, d'une membrane interne, d'un espace périmitochondrial et d'une membrane externe. La membrane mitochondriale externe sépare l'organite de l'hyaloplasme. Habituellement, il a des contours lisses et est fermé de sorte qu'il s'agit d'un sac à membrane.
La membrane externe de l'intérieur est séparée par un espace périmitochondrial d'une largeur d'environ 10 ... 20 nm. La membrane mitochondriale interne limite le contenu interne réel des mitochondries - la matrice. La membrane interne forme de nombreuses protubérances à l'intérieur des mitochondries, qui ont l'apparence de crêtes plates ou de crêtes.
Les crêtes peuvent prendre la forme de plaques (trabéculaires) et de tubules (multivesiculaires à la coupe), et elles sont dirigées longitudinalement ou transversalement par rapport aux mitochondries.
Chaque mitochondrie est remplie d'une matrice qui, sur les micrographies électroniques, semble plus dense que le cytoplasme environnant. La matrice mitochondriale est homogène (homogène), parfois à grains fins, de différentes densités électroniques. Des filaments minces d'une épaisseur d'environ 2 ... 3 nm et des granules d'une taille d'environ 15 ... 20 nm y sont révélés. Les filaments matriciels sont des molécules d'ADN et les petits granules sont des ribosomes mitochondriaux. La matrice contient des enzymes, un ADN cyclique simple brin, des ribosomes mitochondriaux, de nombreux ions Ca 2+.
Le système autonome de synthèse protéique des mitochondries est représenté par des molécules d'ADN sans histone. L'ADN est court, a la forme d'un anneau (cyclique) et contient 37 gènes. Contrairement à l'ADN nucléaire, il ne contient pratiquement aucune séquence nucléotidique non codante. Les caractéristiques de la structure et de l'organisation réunissent l'ADN mitochondrial avec l'ADN des cellules bactériennes. Sur l'ADN mitochondrial, différents types de molécules d'ARN sont synthétisés: informationnel, transfert (transport) et ribosomal. L'ARN d'information des mitochondries n'est pas soumis à un épissage (découpe des sections qui ne portent pas de charge d'information). La petite taille des molécules d'ADN mitochondrial ne peut pas déterminer la synthèse de toutes les protéines mitochondriales. La plupart des protéines mitochondriales sont sous le contrôle génétique du noyau cellulaire et sont synthétisées dans le cytoplasme, car l'ADN mitochondrial est faiblement exprimé et peut fournir la formation d'une partie seulement des enzymes de la chaîne de phosphorylation oxydative. L'ADN mitochondrial code pas plus de dix protéines qui sont localisées dans les membranes et sont des protéines structurelles responsables de l'intégration correcte des complexes protéiques fonctionnels individuels des membranes mitochondriales. Les protéines qui remplissent des fonctions de transport sont également synthétisées. Un tel système de synthèse protéique n'assure pas toutes les fonctions des mitochondries, donc l'autonomie des mitochondries est limitée et relative.
Chez les mammifères, les mitochondries pendant la fécondation ne sont transmises que par l'œuf et le sperme introduit l'ADN du noyau dans le nouvel organisme.
Dans la matrice des mitochondries, des ribosomes se forment qui diffèrent des ribosomes cytoplasmiques. Ils participent à la synthèse d'un certain nombre de protéines mitochondriales non codées par le noyau. Les ribosomes mitochondriaux ont un nombre de sédimentation de 60 (contrairement à cytoplasmique avec un nombre de sédimentation de 80). Le nombre de sédimentation est le taux de dépôt pendant la centrifugation et l'ultracentrifugation. Dans la structure, les ribosomes mitochondriaux sont proches des ribosomes des organismes procaryotes, mais sont plus petits et sont sensibles à certains antibiotiques (chloramphénicol, tétracycline, etc.).
La membrane mitochondriale interne présente un degré élevé de sélectivité dans le transport des substances. Les enzymes de la chaîne de phosphorylation oxydative, les protéines porteuses d'électrons, l'ATP, l'ADP, le pyruvate et d'autres systèmes de transport étroitement adjacents les uns aux autres sont attachés à sa surface interne. En raison de la disposition étroite des enzymes sur la membrane interne, une forte conjugaison (interconnexion) des processus biochimiques est assurée, ce qui augmente le taux et l'efficacité des processus catalytiques.
La microscopie électronique révèle des particules de champignons faisant saillie dans la lumière de la matrice. Ils ont une activité ATP-synthétique (forme l'ATP de l'ADP). Le transport d'électrons a lieu le long de la chaîne respiratoire localisée dans la membrane interne, qui contient quatre grands complexes enzymatiques (cytochromes). Lorsque les électrons traversent la chaîne respiratoire, les ions hydrogène sont pompés de la matrice dans l'espace périmitochondrial, ce qui assure la formation d'un gradient de protons (pompe). L'énergie de ce gradient (différences de concentration de substances et formation de potentiel membranaire) est utilisée pour la synthèse de l'ATP et le transport des métabolites et des ions inorganiques. Les protéines porteuses contenues dans la membrane interne transportent à travers elle les phosphates organiques, l'ATP, l'ADP, les acides aminés, les acides gras, les acides tri- et dicarboxyliques.
La membrane mitochondriale externe est plus perméable aux substances de faible poids moléculaire, car elle contient de nombreux canaux protéiques hydrophiles. Des complexes récepteurs spécifiques sont situés sur la membrane externe, à travers lesquels les protéines de la matrice sont transportées dans l'espace périmitochondrial.
En termes de composition chimique et de propriétés, la membrane externe est proche des autres membranes intracellulaires et du plasmolemme. Il contient des enzymes qui métabolisent les graisses, activent (catalysent) la conversion des amines, l'amino-oxydase. Si les enzymes de la membrane externe restent actives, c'est un indicateur de la sécurité fonctionnelle des mitochondries.
Il existe deux sous-compartiments autonomes dans les mitochondries. L'espace périmitochondrial, ou la chambre mitochondriale externe (sous-compartiment externe), est formé en raison de la pénétration des complexes protéiques de l'hyaloplasme, tandis que le sous-compartiment interne (matrice mitochondriale) est partiellement formé en raison de l'activité synthétique de l'ADN mitochondrial. Le sous-compartiment interne (matrice) contient de l'ADN, de l'ARN et des ribosomes. Il a un niveau élevé d'ions Ca 2+ par rapport à l'hyaloplasme. Les ions hydrogène s'accumulent dans le sous-compartiment externe. L'activité enzymatique des sous-compartiments externe et interne, la composition des protéines sont très différentes. Le sous-compartiment interne a une densité d'électrons plus élevée que l'extérieur.
Des marqueurs mitochondriaux spécifiques sont les enzymes cytochrome oxydase et succinate déshydrogénase, dont la détection permet une caractérisation quantitative des processus énergétiques dans les mitochondries.
La fonction principale des mitochondries - Synthèse ATP. Initialement, dans l'hyaloplasme, les sucres (glucose) sont détruits en acides lactiques et pyruviques (pyruvate) avec la synthèse simultanée d'une petite quantité d'ATP. À la suite de la glycolyse d'une molécule de glucose, deux molécules d'ATP sont utilisées et quatre se forment. Ainsi, seules deux molécules d'ATP constituent l'équilibre positif. Ces processus se produisent sans oxygène (glycolyse anaérobie).
Toutes les étapes ultérieures de la production d'énergie se produisent dans le processus d'oxydation aérobie, qui fournit la synthèse d'une grande quantité d'ATP. Dans ce cas, la matière organique est détruite en CO 2 et en eau. L'oxydation s'accompagne du transfert de protons à leurs accepteurs. Ces réactions sont effectuées en utilisant un certain nombre d'enzymes du cycle de l'acide tricarboxylique, qui sont situées dans la matrice mitochondriale.
Les systèmes de transfert d'électrons et sa phosphorylation associée d'ADP (phosphorylation oxydative) sont intégrés dans les membranes cristallines. Dans ce cas, le transfert d'électrons d'une protéine accepteur d'électrons à une autre a lieu et, enfin, leur liaison à l'oxygène, à la suite de quoi l'eau est formée. Dans le même temps, une partie de l'énergie libérée lors de cette oxydation dans la chaîne de transfert d'électrons est stockée sous forme d'une liaison macroergique lors de la phosphorylation de l'ADP, ce qui conduit à la formation d'un grand nombre de molécules d'ATP - le principal équivalent énergétique intracellulaire. Le processus de phosphorylation oxydative se produit sur les membranes des crêtes mitochondriales en utilisant les protéines de la chaîne d'oxydation et l'enzyme de phosphorylation ADP ATP synthetase située ici. À la suite de la phosphorylation oxydative, 36 molécules d'ATP sont formées à partir d'une molécule de glucose.
Pour certaines hormones et substances présentes dans les mitochondries, il existe des récepteurs spécialisés (affinité). La triiodothyronine accélère normalement l'activité synthétique des mitochondries. L'interleukine-1 et des concentrations élevées de triiodothyronine déconnectent les chaînes de phosphorylation oxydative, provoquent un gonflement mitochondrial, qui s'accompagne d'une augmentation de la formation d'énergie thermique.
Les nouvelles mitochondries se forment par division, constriction ou bourgeonnement. Dans ce dernier cas, des protomitochondries se forment, augmentant progressivement de taille.
Les protomitochondries sont un petit organite avec des membranes externes et internes. La membrane interne n'a pas ou ne contient pas de crêtes sous-développées. Organella se caractérise par de faibles niveaux de phosphorylation aérobie. Avec la formation d'une constriction, le contenu des mitochondries est réparti entre deux nouveaux organites assez gros. Avec n'importe quelle méthode de reproduction, chacune des mitochondries nouvellement formées possède son propre génome.
Les mitochondries plus anciennes sont détruites par autolyse (auto-digestion par la cellule à l'aide de lysosomes) pour former des autolysosomes. À partir de l'autolysosome, un corps résiduel se forme. Avec une digestion complète, le contenu du corps résiduel, composé de substances organiques de faible poids moléculaire, est excrété par exocytose. Avec une digestion incomplète, les restes de mitochondries peuvent s'accumuler dans la cellule sous forme de couches ou de granules avec de la nipofuscine. Les sels de calcium insolubles s'accumulent dans la partie des mitochondries avec la formation de cristaux - calcifications. L'accumulation de produits de dégénérescence mitochondriale peut entraîner une dystrophie cellulaire.
La structure et la fonction des mitochondries est une question assez complexe. La présence d'un organite est caractéristique de presque tous les organismes nucléaires - à la fois pour les autotrophes (plantes capables de photosynthèse) et pour les hétérotrophes, qui sont presque tous des animaux, certaines plantes et des champignons.
Le but principal des mitochondries est l'oxydation des substances organiques et l'utilisation ultérieure de l'énergie libérée à la suite de ce processus. Pour cette raison, les organites ont également un deuxième nom (non officiel) - stations d'énergie cellulaire. Ils sont parfois appelés «plastes de catabolisme».
Qu'est-ce que les mitochondries
Le terme est d'origine grecque. Traduit, ce mot signifie "fil" (mitos), "graine" (chondrion). Les mitochondries sont des organoïdes permanents qui sont d'une grande importance pour le fonctionnement normal des cellules et permettent à tout l'organisme d'exister.

Les «stations» ont une structure interne spécifique, qui varie en fonction de l'état fonctionnel des mitochondries. Leur forme peut être de deux types - ovale ou oblongue. Ce dernier a souvent un aspect ramifié. Le nombre d'organites dans une cellule varie de 150 à 1500.
Un cas particulier est celui des cellules germinales. Une seule organelle spirale est présente dans le sperme, tandis que les gamètes femelles contiennent des centaines de milliers d'autres mitochondries. Dans la cellule, les organites ne sont pas fixés au même endroit, mais peuvent se déplacer le long du cytoplasme, se combiner les uns avec les autres. Leur taille est de 0,5 microns, la longueur peut atteindre 60 microns, tandis que le chiffre minimum est de 7 microns.

Déterminer la taille d'une «centrale électrique» n'est pas une tâche facile. Le fait est que, vu au microscope électronique, seule une partie de l'organite pénètre dans la tranche. Il se trouve que les mitochondries spirales ont plusieurs sections qui peuvent être prises pour des structures séparées et indépendantes.
Seule une image tridimensionnelle permettra de connaître la structure cellulaire exacte et de comprendre si nous parlons de 2 à 5 organoïdes séparés ou de celui avec une forme complexe de mitochondries.
Caractéristiques structurelles
La membrane mitochondriale se compose de deux couches: externe et interne. Ce dernier comprend diverses excroissances et plis qui ont une forme de feuille et tubulaire.
Chaque membrane a une composition chimique spéciale, une certaine quantité de certaines enzymes et un objectif spécifique. L'enveloppe extérieure de l'intérieur est séparée par un espace intermembranaire d'une épaisseur de 10-20 nm.
La structure de l'organite dans la figure avec les légendes semble très claire.

La structure des mitochondries
En regardant le diagramme de structure, la description suivante peut être faite. L'espace visqueux à l'intérieur des mitochondries s'appelle la matrice. Sa composition crée un environnement favorable pour l'écoulement des processus chimiques nécessaires en son sein. Il contient des granules microscopiques qui favorisent les réactions et les processus biochimiques (par exemple, accumulent des ions glycogènes et d'autres substances).
La matrice contient de l'ADN, des coenzymes, des ribosomes, de l'ARN-t, des ions inorganiques. L'ATP synthase et les cytochromes sont situés à la surface de la couche interne de la coquille. Les enzymes contribuent à des processus tels que le cycle de Krebs (CTK), la phosphorylation oxydative, etc.
Ainsi, la tâche principale de l'organoïde est effectuée à la fois par la matrice et par la face intérieure de la coquille.
Fonction mitochondriale
La finalité des «centrales électriques» peut être caractérisée par deux tâches principales:
- production d'énergie: des processus oxydatifs y sont effectués avec la libération ultérieure de molécules d'ATP;
- stockage d'informations génétiques;
- participation à la synthèse d'hormones, d'acides aminés et d'autres structures.
Le processus d'oxydation et de production d'énergie se déroule en plusieurs étapes:


Dessin schématique de la synthèse d'ATP
Il convient de noter: à la suite du cycle de Krebs (cycle de l'acide citrique), les molécules d'ATP ne se forment pas, les molécules sont oxydées et le dioxyde de carbone est libéré. Il s'agit d'une étape intermédiaire entre la glycolyse et la chaîne de transport d'électrons.
Tableau "Fonctions et structure des mitochondries"

Ce qui détermine le nombre de mitochondries dans une cellule
Le nombre actuel d'organoïdes s'accumule près des parties de la cellule où il y a un besoin de ressources énergétiques. En particulier, un grand nombre d'organites sont collectés dans la zone des myofibrilles, qui font partie des cellules musculaires qui assurent leur contraction.

Dans les cellules germinales mâles, les structures sont localisées autour de l'axe du flagelle - on suppose que le besoin d'ATP est dû au mouvement constant de la queue du gamète. La disposition des mitochondries dans les protozoaires, qui utilisent des cils spéciaux pour le mouvement, ressemble à des organites s'accumulant sous la membrane à leur base exactement de la même manière.
Quant aux cellules nerveuses, la localisation des mitochondries est observée à proximité des synapses par lesquelles les signaux du système nerveux sont transmis. Dans les cellules synthétisant des protéines, les organites s'accumulent dans les zones ergastoplasmiques - elles fournissent l'énergie qui fournit ce processus.
Qui a découvert les mitochondries
La structure cellulaire a gagné son nom en 1897-1898 grâce à C. Brand. Otto Wagburg a pu prouver la relation de la respiration cellulaire avec les mitochondries en 1920.
Conclusion
Les mitochondries sont le composant le plus important d'une cellule vivante, agissant comme une station d'énergie qui produit des molécules d'ATP, assurant ainsi les processus d'activité cellulaire.
Le travail des mitochondries est basé sur l'oxydation des composés organiques, entraînant la génération d'un potentiel énergétique.
Membrane extérieure
Membrane intérieure
Matrix m-on, matrice, krista. il a des contours lisses, ne forme ni protubérances ni plis. Il représente environ 7% de la surface de toutes les membranes cellulaires. Son épaisseur est d'environ 7 nm, il n'est connecté à aucune autre membrane du cytoplasme et est fermé à lui-même, il s'agit donc d'un sac à membrane. La membrane externe est séparée de la membrane interne espace intermembranaire environ 10-20 nm de large. La membrane interne (environ 7 nm d'épaisseur) limite le contenu interne réel des mitochondries,
sa matrice ou mitoplasme. Une caractéristique de la membrane interne des mitochondries est leur capacité à former de nombreuses saillies à l'intérieur des mitochondries. Ces saillies ont le plus souvent la forme de crêtes plates ou de crêtes. La distance entre les membranes dans la crista est d'environ 10-20 nm. Souvent, les crêtes peuvent se ramifier ou former des processus en forme de doigts, se plier et ne pas avoir une orientation prononcée. Dans les protozoaires, les algues unicellulaires, dans certaines cellules de plantes et d'animaux supérieurs, les excroissances de la membrane interne ressemblent à des tubes (crêtes tubulaires).
La matrice mitochondriale a une structure homogène à grains fins, des filaments parfois minces collectés dans une boule (environ 2-3 nm) et des granules d'environ 15-20 nm y sont détectés. On sait maintenant que les brins de la matrice mitochondriale sont des molécules d'ADN dans la composition du nucléoïde mitochondrial, et les petits granules sont des ribosomes mitochondriaux.
Fonction mitochondriale
1. Dans les mitochondries, la synthèse d'ATP a lieu (voir. Phosphorylation oxydative)
PH intermembranaire ~ 4, pH matriciel ~ 8 | la teneur en protéines en m: 67% est la matrice, 21% est la découverte de la mine, 6% est à l'intérieur de la mine et 6% est dans l'intermine
Distemper - un seul système mitochondrial
m-na externe: les pores de porine permettent le passage jusqu'à 5 kD | m-na intérieur: puits m imperméable à la cardiolipine pour les ions |
production internationale: groupes d'enzymes phosphorylés nucléotides et sucres nucléotidiques
Intérieur:
matrice: enzymes métaboliques - oxydation des lipides, oxydation des glucides, cycle de l'acide tricarboxylique, cycle de Krebs
Origine bactérienne: l'amibe Pelomyxa palustris ne contient pas d'unité eucaryote, vit en symbiose avec les bactéries aérobies | propre ADN | Processus de type OX
ADN mitochondrial
Division des myochondries
répliqué
en interphase | réplication n'est pas associée à la phase S | pendant le cycle cl, les mitochis sont une fois divisés en deux, formant une constriction, la constriction d'abord à l'intérieur m-pas | ~ 16,5 ko | anneau code pour 2 ARNr 22 ARNt et 13 protéines |
transport de protéines: peptide signal | boucle amphiphile | récepteur de reconnaissance mitochondriale |
Phosphorylation oxydative
Circuit de transfert d'électrons
ATP synthase
dans les cellules hépatiques, m en direct ~ 20 jours de division mitochondriale par formation de constriction
16569 pb \u003d 13 protéines, 22tRNA, 2pRNA | m-na externe lisse (porines - perméabilité des protéines jusqu'à 10 kDa) m-na interne (crista) replié (75% de protéines): transport des protéines porteuses, f-you, composants de la chaîne respiratoire et ATP synthase, cardiolipine) matrice ( enrichi en cycle citrate f-tami) inter 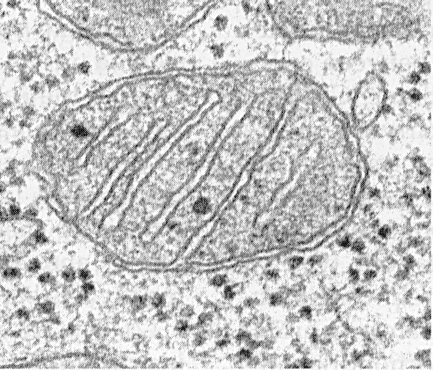
Les mitochondries ou chondriosomes (du grec. Mitos - fil, chondrion - grain, soma - corps) sont des organites granuleux ou filamenteux présents dans le cytoplasme des protozoaires, des plantes et des animaux. Les mitochondries peuvent être observées dans les cellules vivantes, car elles ont une densité assez élevée. Dans les cellules vivantes, les mitochondries peuvent se déplacer, se déplacer, fusionner les unes avec les autres.
Chez différentes espèces, les tailles des mitochondries sont très variables, ainsi que leur forme est variable (Fig.199). Néanmoins, dans la plupart des cellules, l'épaisseur de ces structures est relativement constante (environ 0,5 μm), et la longueur fluctue, atteignant jusqu'à 7-60 μm sous formes filamenteuses.
L'étude de la taille et du nombre de mitochondries n'est pas si simple. Cela est dû au fait que la taille et le nombre de mitochondries visibles sur les coupes ultra-minces ne correspondent pas à la réalité.
Les calculs classiques montrent qu'environ 200 mitochondries sont présentes dans la cellule hépatique. Cela représente plus de 20% du cytoplasme total et environ 30 à 35% de la quantité totale de protéines dans la cellule. La surface de toutes les mitochondries de la cellule hépatique est 4 à 5 fois plus grande que la surface de sa membrane plasmique. La plupart des mitochondries se trouvent dans les ovocytes (environ 300 000) et dans le chaos géant de l'amibe Chaos (jusqu'à 500 000).
Dans les cellules végétales vertes, le nombre de mitochondries est inférieur à celui des cellules animales, car une partie de leurs fonctions peut être assurée par des chloroplastes.
La localisation des mitochondries dans les cellules est différente. Typiquement, les mitochondries s'accumulent près des zones du cytoplasme où il y a un besoin d'ATP formé dans les mitochondries. Ainsi, dans le muscle squelettique, les mitochondries sont situées près des myofibrilles. Dans les spermatozoïdes, les mitochondries forment un boîtier en spirale autour de l'axe du flagelle; cela est probablement dû à la nécessité d'utiliser l'ATP pour le mouvement de la queue du sperme. De même, dans les protozoaires et dans d'autres cellules équipées de cils, les mitochondries sont localisées directement sous la membrane cellulaire à la base des cils, ce qui nécessite que l'ATP fonctionne. Dans les axones des cellules nerveuses, les mitochondries sont situées près des synapses, où se produit le processus de transmission d'une impulsion nerveuse. Dans les cellules sécrétoires qui synthétisent de grandes quantités de protéines, les mitochondries sont étroitement associées aux zones ergastoplasmes; ils fournissent probablement de l'ATP pour l'activation des acides aminés et la synthèse des protéines sur les ribosomes.
Ultrastructure des mitochondries.
Les mitochondries, quelle que soit leur taille ou leur forme, ont une structure universelle, leur ultrastructure est uniforme. Les mitochondries sont limitées à deux membranes (Fig.205). La membrane mitochondriale externe la sépare de l'hyaloplasme, elle a des contours lisses, ne forme pas de protubérances ou de plis, et l'épaisseur est d'environ 7 nm. Il représente environ 7% de la surface de toutes les membranes cellulaires. La membrane n'est reliée à aucune autre membrane cytoplasmique, est fermée sur elle-même et est un sac à membrane. La membrane externe de l'intérieur est séparée par un espace intermembranaire d'une largeur d'environ 10-20 nm. La membrane interne (environ 7 nm d'épaisseur) limite le contenu interne réel des mitochondries, de sa matrice ou de son mitoplasme. La membrane mitochondriale interne forme de nombreuses intrusions à l'intérieur des mitochondries. Ces saillies ont le plus souvent la forme de crêtes plates ou de crêtes.
La surface totale de la membrane mitochondriale interne dans la cellule hépatique est d'environ un tiers de la surface de toutes les membranes cellulaires. Les mitochondries des cellules du muscle cardiaque contiennent trois fois plus de crêtes que les mitochondries hépatiques, ce qui reflète les différences dans les charges fonctionnelles des mitochondries des différentes cellules. La distance entre les membranes dans la crista est d'environ 10-20 nm.
Les crêtes mitochondriales s'étendant de la membrane interne et s'étendant vers la matrice ne bloquent pas complètement la cavité mitochondriale et ne violent pas la continuité de la matrice qui la remplit.
L'orientation des crêtes par rapport à l'axe long des mitochondries est différente pour différentes cellules. L'orientation peut être perpendiculaire (foie, cellules rénales); dans le muscle cardiaque, une disposition longitudinale des crêtes est observée. Crista peut se ramifier ou former des processus en forme de doigts, se plier et ne pas avoir une orientation prononcée (Fig. 208). Dans les protozoaires, les algues unicellulaires, dans certaines cellules de plantes et d'animaux supérieurs, les excroissances de la membrane interne ressemblent à des tubes (crêtes tubulaires).
La matrice mitochondriale a une structure homogène à grains fins dans laquelle les molécules d'ADN sont détectées sous la forme de filaments minces collectés dans une boule (environ 2-3 nm) et les ribosomes mitochondriaux sous la forme de granules d'environ 15-20 nm. Les dépôts de sels de magnésium et de calcium dans la matrice forment de gros granules denses (20-40 nm).
Les fonctions des mitochondries.
Les mitochondries effectuent la synthèse de l'ATP, qui se produit à la suite de l'oxydation des substrats organiques et de la phosphorylation de l'ADP.
Les étapes initiales de l'oxydation des glucides sont appelées oxydation anaérobie, ou glycolyse et se produisent dans l'hyaloplasme et ne nécessitent pas d'oxygène. Le substrat d'oxydation pendant la production d'énergie anaérobie sont les hexoses et principalement le glucose; certaines bactéries ont la capacité d'extraire de l'énergie en oxydant les pentoses, les acides gras ou les acides aminés.
Dans le glucose, la quantité d'énergie potentielle contenue dans les liaisons entre les atomes C, H et O est d'environ 680 kcal pour 1 mole (c'est-à-dire 180 g de glucose).
Dans une cellule vivante, cette énorme quantité d'énergie est libérée sous la forme d'un processus par étapes, contrôlé par un certain nombre d'enzymes oxydantes, et n'est pas associée à la transition de l'énergie des liaisons chimiques en chaleur, comme lors de la combustion, mais passe dans une liaison macroénergétique dans les molécules d'ATP, qui sont synthétisées en utilisant l'énergie libérée de ADP et phosphate.
Les trioses formées à la suite de la glycolyse, principalement l'acide pyruvique, sont impliquées dans les mitochondries dans l'oxydation ultérieure. Dans ce cas, l'énergie de clivage de toutes les liaisons chimiques est utilisée, ce qui conduit à la libération de CO 2, à la consommation d'oxygène et à la synthèse d'une grande quantité d'ATP. Ces processus sont associés au cycle d'oxydation des acides tricarboxyliques et à la chaîne respiratoire du transfert d'électrons, où se produisent la phosphorylation de l'ADP et la synthèse du «carburant» cellulaire et des molécules d'ATP (Fig. 209).
Dans le cycle de l'acide tricarboxylique (cycle de Krebs ou cycle de l'acide citrique), le pyruvate formé à la suite de la glycolyse perd d'abord la molécule de CO 2 et, étant oxydé en acétate (un composé à deux carbones), se combine avec la coenzyme A. Ensuite, l'acétyl-coenzyme A se combine avec l'oxalacétate (un composé à quatre carbones), forme du citrate à six carbones (acide citrique). Ensuite, il y a un cycle d'oxydation de ce composé à six carbones en oxalacétate à quatre carbones, se liant à nouveau à l'acétyl coenzyme A, puis le cycle se répète. Au cours de cette oxydation, deux molécules de CO 2 sont libérées et les électrons libérés lors de l'oxydation sont transférés aux molécules de coenzyme accepteur (NAD-nicotinamide adénine dinucléotide), qui les impliquent davantage dans la chaîne de transfert d'électrons. Par conséquent, dans le cycle des acides tricarboxyliques, il n'y a pas de synthèse d'ATP elle-même, mais il y a une oxydation des molécules, un transfert d'électrons vers les accepteurs et une émission de CO 2. Tous les événements décrits ci-dessus dans les mitochondries se produisent dans leur matrice.
L'oxydation du substrat initial entraîne la libération de CO 2 et d'eau, mais l'énergie thermique n'est pas libérée, comme lors de la combustion, mais des molécules d'ATP se forment. Ils sont synthétisés par un autre groupe de protéines qui ne sont pas directement liées à l'oxydation. Dans les membranes mitochondriales internes, de grands complexes protéiques, des enzymes, des ATP synthétases sont situés à la surface des membranes qui regardent dans la matrice. Au microscope électronique, ils sont visibles sous la forme de corps dits «champignons», qui tapissent complètement la surface des membranes, en regardant dans la matrice. Les corps ont une jambe et une tête, d'un diamètre de 8 à 9 nm. Par conséquent, les enzymes de la chaîne oxydante et les enzymes de synthèse de l'ATP sont localisées dans les membranes mitochondriales internes (Fig. 201b).
La chaîne respiratoire est le principal système de conversion d'énergie dans les mitochondries. Ici, l'oxydation et la restauration séquentielles des éléments de la chaîne respiratoire se produisent, à la suite desquelles l'énergie est libérée en petites portions. En raison de cette énergie, l'ATP est formé d'ADP et de phosphate à trois points de la chaîne. Par conséquent, ils disent que l'oxydation (transfert d'électrons) est associée à la phosphorylation (ADP + Fn → ATP, c'est-à-dire que la phosphorylation oxydative se produit.
L'énergie libérée lors du transport d'électrons est stockée sous forme d'un gradient de protons sur la membrane. Il s'est avéré que lors du transfert d'électrons dans la membrane mitochondriale, chaque complexe de la chaîne respiratoire dirige l'énergie libre d'oxydation pour déplacer les protons (charges positives) à travers la membrane, de la matrice à l'espace intermembranaire, ce qui conduit à la formation de différences potentielles sur la membrane: des charges positives prévalent dans l'espace intermembranaire, et négatif - du côté de la matrice mitochondriale. Lorsque la différence de potentiel (220 mV) est atteinte, le complexe protéique de l'ATP synthétase commence à transporter les protons vers la matrice, tandis qu'il transforme une forme d'énergie en une autre: il forme l'ATP à partir de l'ADP et du phosphate inorganique. Il s'agit de la combinaison de processus oxydatifs et de processus synthétiques, avec phosphorylation d'ADP. Alors que les substrats s'oxydent, tandis que les protons sont pompés à travers la membrane mitochondriale interne, la synthèse d'ATP associée à cela se produit, c'est-à-dire il se produit une phosphorylation oxydative.
Ces deux processus peuvent être séparés. Dans ce cas, le transfert d'électrons se poursuit, ainsi que l'oxydation du substrat, mais la synthèse d'ATP ne se produit pas. Dans ce cas, l'énergie libérée lors de l'oxydation est convertie en énergie thermique.
Phosphorylation oxydative chez les bactéries.
Dans les cellules procaryotes capables de phosphorylation oxydative, les éléments du cycle de l'acide tricarboxylique sont localisés directement dans le cytoplasme, et la chaîne respiratoire et les enzymes de phosphorylation sont associées à la membrane cellulaire, avec ses saillies faisant saillie à l'intérieur du cytoplasme, avec les soi-disant mésosomes (Fig.212). Il convient de noter que ces mésosomes bactériens peuvent être associés non seulement aux processus de respiration aérobie, mais aussi chez certaines espèces, participent à la division cellulaire, au processus de distribution d'ADN dans les nouvelles cellules, à la formation de la paroi cellulaire, etc.
Sur la membrane plasmique des mésosomes de certaines bactéries, des processus conjugués d'oxydation et de synthèse d'ATP sont effectués. Des particules sphériques similaires à celles trouvées dans les mitochondries des cellules eucaryotes ont été trouvées au microscope électronique dans les fractions des membranes plasmiques des bactéries. Ainsi, dans les cellules bactériennes capables de phosphorylation oxydative, la membrane plasmique joue un rôle similaire à la membrane mitochondriale interne des cellules eucaryotes.
Une augmentation du nombre de mitochondries.
Les mitochondries peuvent augmenter leur nombre, en particulier avec la division cellulaire ou avec une augmentation de la charge fonctionnelle de la cellule. Il y a un renouvellement constant des mitochondries. Par exemple, dans le foie, la durée de vie moyenne des mitochondries est d'environ 10 jours.
Une augmentation du nombre de mitochondries se produit par la croissance et la division des mitochondries précédentes. Cette hypothèse a été émise pour la première fois par Altman (1893), qui a décrit les mitochondries sous le terme de «bioblastes». Il est possible d'observer la division in vivo, la fragmentation des mitochondries longues en plus courtes par constriction, qui ressemble à la méthode binaire de division des bactéries.
Une réelle augmentation du nombre de mitochondries par division a été établie lors de l'étude du comportement des mitochondries dans les cellules vivantes de culture tissulaire. Au cours du cycle cellulaire, les mitochondries atteignent plusieurs microns, puis se fragmentent, se divisent en corps plus petits.
Les mitochondries peuvent fusionner et se multiplier selon le principe: les mitochondries des mitochondries.
Autoproduction de mitochondries.
Les organites à deux membranes ont un système complet d'auto-reproduction. Dans les mitochondries et les plastes, il y a de l'ADN sur lequel sont synthétisés des informations, des transferts et des ARN ribosomaux et des ribosomes, qui effectuent la synthèse des protéines mitochondriales et des plastes. Cependant, ces systèmes, bien qu'autonomes, sont limités dans leurs capacités.
L'ADN dans les mitochondries est une molécule cyclique sans histones et ressemble ainsi aux chromosomes bactériens. Leur taille est d'environ 7 microns, 16 à 19 000 paires de nucléotides d'ADN entrent dans une molécule cyclique de mitochondries animales. Chez l'homme, l'ADN mitochondrial contient 16,5 mille n.p., il est complètement déchiffré. Il a été constaté que l'ADN mitochondral de divers objets est très homogène, leur différence réside uniquement dans l'ampleur des introns et des sites non transcrits. Tous les ADN mitochondriaux sont des copies multiples assemblées en groupes, grappes. Ainsi, dans une mitochondrie du foie de rat peut contenir de 1 à 50 molécules d'ADN cycliques. La quantité totale d'ADN mitochondrial par cellule est d'environ un pour cent. La synthèse de l'ADN mitochondrial n'est pas liée à la synthèse de l'ADN dans le noyau.
Comme dans les bactéries, l'ADN mitochondral est collecté dans une zone distincte - le nucléoïde, sa taille est d'environ 0,4 microns de diamètre. Dans les longues mitochondries, il peut y avoir de 1 à 10 nucléoïdes. Lors de la division d'une longue mitochondrie, une section contenant un nucléoïde en est séparée (similaire à la division binaire des bactéries). La quantité d'ADN dans les nucléoïdes mitochondriaux individuels peut fluctuer 10 fois selon le type de cellules.
Dans certaines cultures, dans les cellules de 6 à 60% des mitochondries n'ont pas de nucléoïde, ce qui peut s'expliquer par le fait que la division de ces organites est plus vraisemblablement associée à la fragmentation plutôt qu'à la distribution des nucléoïdes.
Comme déjà mentionné, les mitochondries peuvent à la fois se diviser et se fusionner. Lorsque les mitochondries fusionnent les unes avec les autres, un échange de leurs composants internes peut se produire.
Il est important de souligner que l'ARNr et les ribosomes des mitochondries et du cytoplasme sont très différents. Si des ribosomes des années 80 se trouvent dans le cytoplasme, alors les ribosomes mitochondriaux des cellules végétales appartiennent aux ribosomes des années 70 (se composent de sous-unités des années 30 et 50, contiennent des ARN 16 et 23 caractéristiques des cellules procaryotes), et des ribosomes plus petits se trouvent dans les mitochondries des cellules animales (environ 50).
Les ARN ribosomiques mitochondriaux sont synthétisés sur l'ADN mitochondrial. Dans le mitoplasme sur les ribosomes se trouve la synthèse des protéines. Il s'arrête, contrairement à la synthèse sur les ribosomes cytoplasmiques, sous l'action de l'antibiotique chloramphénicol, qui inhibe la synthèse des protéines dans les bactéries.
Sur le génome mitochondrial, 22 ARN de transport sont synthétisés. Le code triplet du système synthétique mitochondrial est différent de celui utilisé dans l'hyaloplasme. Malgré la présence apparemment de tous les composants nécessaires à la synthèse des protéines, les petites molécules d'ADN mitochondrial ne peuvent pas coder toutes les protéines mitochondriales, seulement une petite fraction d'entre elles. Donc, l'ADN de 15 mille n.p. peut coder pour des protéines d'un poids moléculaire total d'environ 6 x 10 5. Dans le même temps, le poids moléculaire total des protéines des particules de l'ensemble respiratoire complet des mitochondries atteint une valeur d'environ 2x10 6.
Étant donné qu'en plus des protéines de phosphorylation oxydative, les mitochondries comprennent également les enzymes du cycle de l'acide tricarboxylique, les enzymes de synthèse d'ADN et d'ARN, les enzymes d'activation des acides aminés et d'autres protéines, on peut voir que pour coder ces nombreuses protéines et ARNr et ARNt, la quantité d'informations génétiques dans la courte molécule d'ADN mitochondrial n'est clairement pas suffisante. Le décodage de la séquence nucléotidique de l'ADN mitochondrial humain a montré qu'il ne code que 2 ARN ribosomiques, 22 ARN de transfert et un total de 13 chaînes polypeptidiques différentes.
Il a maintenant été prouvé que la plupart des protéines mitochondriales sont sous contrôle génétique à partir du noyau cellulaire et sont synthétisées en dehors des mitochondries. La plupart des protéines mitochondriales sont synthétisées sur des ribosomes dans le cytosol. Ces protéines ont des séquences signal spéciales reconnues par les récepteurs de la membrane mitochondriale externe. Ces protéines peuvent y être insérées (voir l'analogie avec la membrane peroxysome), puis se déplacer vers la membrane interne. Ce transfert se produit aux points de contact des membranes externe et interne où un tel transport est noté. La plupart des lipides mitochondriaux sont également synthétisés dans le cytoplasme.
Tout cela indique l'origine endosymbiotique des mitochondries, que les mitochondries sont des organismes tels que les bactéries, qui sont en symbiose avec une cellule eucaryote.
Chondriom.
La totalité de toutes les mitochondries dans une cellule est appelée chondriome. Cela peut être différent selon le type de cellules. Dans de nombreuses cellules, les chondriomes sont constitués de nombreuses mitochondries dispersées uniformément situées dans tout le cytoplasme ou localisées en groupes aux sites de dépenses d'ATP intenses. Dans ces deux cas, les mitochondries fonctionnent seules, leur travail coopératif, éventuellement coordonné par certains signaux du cytoplasme. Il existe un type de chondriome complètement différent, lorsqu'au lieu de petites mitochondries dispersées uniques, une mitochondrie géante ramifiée est située dans la cellule.
Ces mitochondries se trouvent dans les algues vertes unicellulaires (par exemple, Chlorella). Ils forment un réseau mitochondrial complexe ou réticulum mitochondrial (Reticulum miyochondriale). Selon la théorie chimio-osmotique, la signification biologique de l'apparition d'une telle structure mitochondriale ramifiée géante, réunie par ses membranes externe et interne, est que la synthèse d'ATP peut se produire en tout point de la surface de la membrane interne d'une telle mitochondrie ramifiée, qui ira n'importe où dans le cytoplasme, où Il y a un besoin pour cela.
Dans le cas de mitochondries géantes ramifiées, en tout point de la membrane interne, un potentiel suffisant pour démarrer la synthèse de l'ATP peut s'accumuler. De ces positions, le réticulum mitochondrial est, pour ainsi dire, un conducteur électrique, un câble reliant les points éloignés d'un tel système. Le réticulum mitochondrial s'est révélé très utile non seulement pour les petites cellules mobiles, telles que la chlorelle, mais également pour les unités structurales plus grandes telles que, par exemple, les myofibrilles dans le muscle squelettique.
On sait que le muscle squelettique est constitué d'une masse de fibres musculaires, symplastes contenant de nombreux noyaux. La longueur de ces fibres musculaires atteint 40 microns, avec une épaisseur de 0,1 microns - c'est une structure gigantesque contenant un grand nombre de myofibrilles, qui se contractent toutes simultanément, de manière synchrone. Pour la contraction, une grande quantité d'ATP, que les mitochondries fournissent au niveau des disques z, est délivrée à chaque unité de contraction, à la myofibril. Sur les coupes longitudinales ultraminces des muscles squelettiques au microscope électronique, de nombreuses petites coupes arrondies de mitochondries situées au voisinage des sarcomes sont visibles. Les mitochondries musculaires ne sont pas de petites boules ou tiges, mais plutôt des structures en forme d'araignée, dont les processus se ramifient et s'étendent sur de longues distances, parfois sur tout le diamètre de la fibre musculaire.
En même temps, une ramification mitochondriale entoure chaque myofibril dans la fibre musculaire, leur fournissant l'ATP nécessaire à la contraction musculaire. Par conséquent, dans le plan du disque z, les mitochondries représentent un réticulum mitochondrial typique. Une telle couche ou plancher du réticulum mitochondrial est répété deux fois pour chaque sarcomère, et la fibre musculaire entière a des milliers de couches transversales de «plancher» du réticulum mitochondrial. Entre les «planchers» le long des myofibrilles se trouvent des mitochondries filamenteuses reliant ces couches mitochondriales. Ainsi, une image en trois dimensions du réticulum mitochondrial, traversant tout le volume de fibre musculaire, a été créée.
Il a en outre été établi qu'entre les branches du réticulum mitochondrial et les mitochondries longitudinales filiformes, il existe des composés ou contacts intermitochondriaux spéciaux (MMC). Ils sont formés par des membranes mitochondriales externes étroitement adjacentes de mitochondries en contact, l'espace intermembranaire et les membranes dans cette zone ont une densité électronique accrue. Grâce à ces formations spéciales, une union fonctionnelle des mitochondries et des réticulums mitochondriaux voisins a lieu en un seul système énergétique coopératif. Toutes les myofibrilles dans la fibre musculaire sont réduites de manière synchrone sur toute leur longueur.Par conséquent, l'apport d'ATP sur n'importe quelle partie de cette machine complexe devrait également se produire de manière synchrone, et cela ne peut se produire que si un grand nombre de conducteurs mitochondriaux branchés sont connectés les uns aux autres. en utilisant des contacts.
Le fait que les contacts intermitochodiaux (MMK) soient impliqués dans l'association énergétique des mitochondries les uns avec les autres était possible sur les cardiomyocytes, les cellules du muscle cardiaque.
Les chondriomes des cellules du muscle cardiaque ne forment pas de structures ramifiées, mais sont représentés par de nombreuses petites mitochondries allongées situées sans ordre spécial entre les myofibrilles. Cependant, toutes les mitochondries voisines se rejoignent en utilisant des contacts mitochondriaux du même type que dans le muscle squelettique, seul leur nombre est très grand: en moyenne, il y a 2-3 MMC par mitochondrie qui lient les mitochondries en une seule chaîne, où chaque maillon a (Streptio mitochondriale) est une mitochondrie distincte.
Il s'est avéré que des contacts intermitochondriaux (MMC), en tant que structure obligatoire des cellules cardiaques, ont été trouvés dans les cardiomyocytes des ventricules et des oreillettes de tous les vertébrés: mammifères, oiseaux, reptiles, amphibiens et poissons osseux. De plus, des MMC ont été trouvées (mais dans une moindre mesure) dans les cellules cardiaques de certains insectes et mollusques.
Le nombre de MMK dans les cardiomyocytes varie en fonction de la charge fonctionnelle sur le cœur. Le nombre de MMC augmente avec l'augmentation de l'activité physique des animaux et, inversement, lorsque la charge sur le muscle cardiaque diminue, une forte diminution du nombre de MMC se produit.







